Kimberella
Kimberella est un genre éteint d'organismes à symétrie bilatérale, connu uniquement dans les roches de la période géologique de l'Édiacarien. Deux espèces ont été décrites, Kimberella quadrata et Kimberella persii, pour ces organismes ressemblant à des limaces qui se nourrissaient en éraflant la surface microbienne sur laquelle ils vivaient, d'une manière similaire aux mollusques, bien que leur appartenance à cet embranchement soit controversée.

| Règne | Animalia |
|---|---|
| Sous-règne | Eumetazoa |
| — non classé — | Bilateria |
| Embranchement | Mollusca ? |
Espèces de rang inférieur
- †Kimberella quadrata Glaessner & Wade, 1966
- †Kimberella persii Vaziri, Majidifard & Laflamme, 2018
Les premiers spécimens ont été découverts dans les collines Ediacara en Australie[1], et par la suite de nombreux individus furent trouvés dans la région de la mer Blanche en Russie, datés de 558 à 555 millions d'années[2]. Comme pour de nombreux fossiles de cette époque, les liens de Kimberella à d'autres organismes dans la chaîne évolutive sont vivement débattus. Les paléontologues ont initialement classé ce genre comme un type de méduses, mais depuis 1997, ses caractéristiques anatomiques et son association avec des éraflures ressemblant à celles produites par une radula ont été interprétées comme des signes pour le classer dans les mollusques. Bien que certains paléontologues contestent cette classification, il est généralement reconnu comme étant au moins un bilatérien.
Répartition géographique
Kimberella a été trouvé dans les collines Ediacara en Australie-Méridionale, dans la Formation Ust' Pinega dans la région de la mer Blanche en Russie et dans le centre de l'Iran[3]. Les fossiles de la mer Blanche sont souvent associés aux « animaux » de la faune de l'Édiacarien Tribrachidium et Dickinsonia, à des traces fossiles formant des sentiers sinueux, peut-être faits par Kimberella lui-même, et à des algues. Les dépôts de la mer Blanche ont été datés par radiométrie entre 555,3 ± 0,3 millions d'années (Ma) et 558 Ma à partir du rapport uranium-plomb dans des zircons trouvés dans des couches de cendres volcaniques prises en sandwich entre les couches fossilifères à Kimberella[2]. Des fossiles de Kimberella ont par ailleurs été retrouvés dans des couches plus anciennes et plus jeunes que celles de cet intervalle[4]. Les fossiles provenant des collines Ediacara n'ont pas été datés avec précision. L'espèce Kimberella persii a été retrouvée en Iran dans les séries géologiques de Kushk, datées de l'Ediacarien terminal (~545-541 Ma)[3].
Description
Plus de 1 000 spécimens, représentant des individus à tous les stades de maturité, ont été trouvés dans la région de la mer Blanche, à la base de couches de grès à grains fins[4] - [5]. Le nombre important de spécimens retrouvés, la finesse des grains, et la variété de circonstances dans lesquelles les spécimens ont été préservés, fournissent des informations détaillées sur la forme externe de Kimberella, son anatomie interne, et ses modes de locomotion et d'alimentation[4].
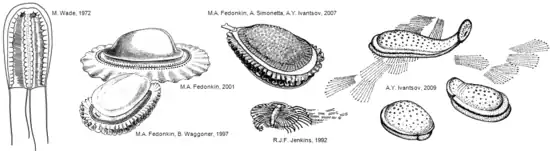
Tous les fossiles sont de forme ovale. Des spécimens allongés illustrent le fait que cet organisme était capable de s'étirer dans une direction antéro-postérieure[5], peut-être jusqu'à un facteur deux. Le seul type de symétrie visible sur les spécimens de la mer Blanche est bilatéral ; il n'y aucun signe d'un des types de symétrie radiale que l'on retrouve dans les cnidaires, embranchement comprenant les méduses, les anémones de mer et les hydrozoaires. Les fossiles australiens ont été initialement décrits comme un type de méduses, mais cela est incompatible avec la symétrie bilatérale présente dans les fossiles. Les fossiles de la mer Blanche et les sédiments environnants montrent que Kimberella vivait sur les fonds marins[6].
Kimberella possède un tégument dorsal qui a été décrit comme une « coquille » non minéralisée. Pour les gros spécimens celui-ci atteint jusqu'à 15 cm de long, 5 à 7 cm de large et 3 à 4 cm de haut ; pour les plus petits spécimens, le tégument n'est que d'environ 2 à 3 mm de long[4]. La coquille est rigide mais flexible, et ne semble pas avoir été minéralisée, durcissant à mesure que celle-ci devenait plus large (et sans doute plus épaisse) pour les individus les plus matures. La déformation observée sur des spécimens allongés et pliés illustre le fait que la coquille était très malléable ; plutôt qu'un tégument unique, il s'agissait peut-être d'une agrégation de sclérites[5]. À son point le plus élevé se trouve une structure en forme de capuchon, que l'on pense être la face avant[6] - [4]. Sur certains échantillons fossiles, la surface interne de la coquille porte des rayures s'étendant sur la largeur de l'individu ; celles-ci pourraient représenter les emplacements de fixation des muscles[4]. Des bandes similaires autour du bord de la coquille pourraient être associées aux muscles de rétractation du pied dans la coquille[4].
L'axe principal de l'organisme est marqué par une crête surélevée et l'axe médian est légèrement bossu. Le corps de Kimberella n'a pas de segmentation visible, mais possède une série de « modules » répétés. Chaque module comprend une bande complètement développée de muscles dorso-ventraux qui vont au haut vers le « pied » unique, large et musclé, et de plus petits muscles transversaux de part et d'autre sur la face inférieure du corps. La combinaison des bandes dorso-ventrales et transversales permettait à Kimberella de se déplacer par ondulation du pied[6] - [4].

Le corps de Kimberella quadrata possède une frange en forme de collerette qui pourrait faire partie du système respiratoire de l'animal, remplissant une fonction similaire à celle des branchies. Le fait que la frange s'étend bien au-delà de la coquille peut indiquer que ces « branchies » étaient inefficaces et avaient besoin d'une grande surface, ou bien qu'il n'existait pas de prédateur redoutable pour Kimberella et que la fonction principale de la coquille était de fournir une plate-forme pour les muscles[4]. Cette collerette est absente chez la seconde espèce décrite, Kimberella persii[3].
Écologie
Kimberella vivait dans des eaux peu profondes (jusqu'à plusieurs dizaines de mètres de profondeur) et partageait des fonds marins calmes et bien oxygénés avec des organismes photosynthétiques et des tapis microbiens. Les assemblages à Kimberella comportent des fossiles de Yorgia, Dickinsonia, Tribrachidium et Charniodiscus, suggérant qu'il vivait aux côtés de ces organismes[4].
Kimberella broutait probablement le tapis microbien, mais il ne peut être exclu qu'il avait une prédation sélective. Il reculait lorsqu'il mangeait. La trace ainsi créée était détruite à la suite du broutage[4]. Des sillons en forme d'éventail, rayonnant à partir de l'organisme, sont souvent trouvés. Ceux-ci indiquent que l'animal restait en un seul endroit et ratissait la surface microbienne vers lui, par un mouvement d'extension de sa « tête », qui portait un certain nombre de « dents » régulièrement espacées[5].
Les eaux dans lesquelles Kimberella habitait étaient parfois perturbées par des courants charriant du sable, causés lorsque les sédiments étaient battus par des tempêtes ou par des libérations d'eaux de fonte, recouvrant les créatures ; apparemment les individus n'étaient pas assez rapides pour aller plus vite que les courants. En réaction à ce stress, les organismes semblent avoir rétracté leurs parties molles dans leur coquille. Certains organismes ont survécu à ces courants et ont tenté de creuser dans le sable déposé sur eux. Des tentatives infructueuses peuvent être vues là où des individus jeunes ont été fossilisés à la fin d'une galerie de quelques centimètres de long[4].
Préservation
Les fossibles de Kimberella sont généralement conservés sur les couches riches en argile et sous une couche de sable[5]. Tous les fossiles sont conservés sous forme de dépressions dans les bases des couches, ce qui implique que l'organisme, bien que non minéralisé, était assez solide pour résister à l'écrasement provoqué par les sédiments accumulés au-dessus de lui. Lorsque les parties molles de l'organisme se décomposaient, les boues molles en dessous auraient été pressées vers le haut dans la coquille, préservant la forme de l'animal[4].
La préservation de la plupart des spécimens a été rendu possible par la sédimentation rapide qui a vite isolé les organismes de l'eau de mer. Elle pourrait avoir été améliorée par la décomposition de l'organisme, aidant les sédiments sus-jacents à minéraliser et à durcir[4]. Il a été avancé qu'une trace de mucus produit par l'organisme peut avoir aidé à la préservation[4], mais des expériences suggèrent que le mucus se désintègre trop facilement pour pouvoir jouer un rôle dans la sédimentation[7].
Classification
La très grande majorité des fossiles de Kimberella trouvés a été attribuée à l'espèce Kimberalla quadrata, décrite en 1966. En 2018, une seconde espèce, Kimberella persii, a été décrite à partir de 9 spécimens découverts en Iran[3]. Les premiers spécimens ont été découverts en Australie en 1959. Ils ont été classés initialement comme des méduses par Martin Glaessner et Mary Wade en 1966[1], puis comme des cubozoaires (méduse-boîte) par Wade en 1972[8], interprétation restée populaire jusqu'à ce que les fossiles de la mer Blanche soient découverts, ce qui a favorisé leur réinterprétation[4]. Les recherches menées sur ces spécimens par Mikhail Fedonkin, initialement en collaboration avec Benjamin M. Waggoner en 1997[6], ont conduit à ce que Kimberalla soit reconnu, de manière bien établie, comme le plus ancien organisme triploblastique (bilératien), et non comme une méduse[9].

Jusqu'à présent, les fossiles retrouvés de Kimberella ne montrent aucun indice d'une radula, sorte de langue munie de dents chitineuses, qui est la caractéristique de diagnose des mollusques modernes, à l'exception des bivalves. Comme les radulas sont très rarement conservées dans les mollusques fossiles, leur absence ne signifie pas nécessairement que K. quadrata n'en était pas dotée. Les roches dans le voisinage immédiat des fossiles de Kimberella présentent des rayures très semblables à celles faites par la radula des mollusques lorsqu'ils broutent sur les tapis microbiens. De telles traces fossiles sont appelées Radulichnus (en) et ont été interprétées comme des preuves indirectes de l'existence d'une radula. En conjonction avec la présence d'une coquille univalve, cela a été pris pour indiquer que Kimberella était un mollusque ou très étroitement lié à ceux-ci[6]. En 2001 et 2007, Fedonkin a suggéré que l'appareil d'alimentation pourrait être un proboscis avec des organes en forme de crochet à son extrémité[4]. Cet appareil d'alimentation semble différer de manière significative de la radula typique des mollusques, et démontrerait que Kimberella est au mieux un représentant du groupe souche des mollusques[11].
Cependant, certains scientifiques, sceptiques quant à cette classification, estiment que les preuves disponibles ne suffisent pas à identifier de manière fiable Kimberella comme un mollusque ou « presque-mollusque », considérant qu'il est présomptueux de l'appeler rien de plus qu'un « possible » mollusque, ou même juste un « bilatérien probable »[12]. Nicholas J. Butterfield fait valoir que l'association de Kimberella avec les marques Radulichnus n'est pas une preuve solide qu'il s'agissait d'un mollusque, d'autres groupes d'organismes possédant des structures capables de faire les traces similaires[12] - [13].
Importance théorique
La majorité des animaux plus complexes que les méduses et autres cnidaires sont divisés en deux groupes : les protostomes et les deutérostomes[9]. Les caractéristiques d'organismes ressemblant aux mollusques comme Kimberella suggèrent fortement que c'était un membre des protostomes[6] - [4]. Dans affirmative, cela signifie que les lignées des protostomes et des deutérostomes ont dû se diviser un certain temps avant l'apparition de Kimberella, il y a au moins 558 millions d'années, et donc bien avant le début du Cambrien il y a 541 millions d'années. Même si ce n'est pas un protostomien, il est largement accepté comme un membre du clade plus large des bilatériens. Depuis que des fossiles de cnidaires à l'allure plutôt moderne ont été retrouvés dans le lagerstätte de Doushantuo, les lignées des cnidaires et des bilatériens auraient divergé il y a bien plus de 580 millions d'années[9].
Références
- (en) M. F. Glaessner et M. Wade, « The late Precambrian fossils from Ediacara, South Australia », Palaeontology, vol. 9, no 4, , p. 599 (lire en ligne [PDF]).
- (en) M. W. Martin et D. V. Grazhdankin, « Age of Neoproterozoic Bilaterian Body and Trace Fossils, White Sea, Russia: Implications for Metazoan Evolution », Science, vol. 288, no 5467, , p. 841-845 (PMID 10797002, DOI 10.1126/science.288.5467.841).
- (en) Seyed Hamid Vaziri, Mahmoud Reza Majidifard et Marc Laflamme, « Diverse Assemblage of Ediacaran fossils from Central Iran », Scientific Reports, vol. 8, no 5060, (DOI 10.1038/s41598-018-23442-y).
- (en) M. A. Fedonkin, A. Simonetta et A. Y. Ivantsov, « New data on Kimberella, the Vendian mollusc-like organism (White Sea region, Russia): palaeoecological and evolutionary implications », dans P. Vickers-Rich et P. Komarower, The rise and fall of the Ediacaran biota, vol. 286, London, Geological Society, coll. « Geological Society special publication », (ISBN 9781862392335, DOI 10.1144/SP286.12, lire en ligne).
- (en) A. Yu. Ivantsov, « New reconstruction of Kimberella, problematic Vendian metazoan », Paleontological Journal, vol. 43, no 6, , p. 601-611 (DOI 10.1134/S003103010906001X, résumé).
- (en) M. A. Fedonkin et B. M. Waggoner, « The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism », Nature, vol. 388, no 6645, , p. 868-871 (DOI 10.1038/42242, Bibcode 1997Natur.388.868F).
- (en) P. R. Getty, « Production and Preserving Climactichnites », dans The Geological Society of America. 2006 Philadelphia Annual Meeting, vol. 38, (lire en ligne), chap. 7, p. 475
- (en) M. Wade, « Hydrozoa and Scyphozoa and other medusoids from the Precambrian Ediacara fauna, South Australia », Palaeontology, vol. 15, , p. 197–225 (lire en ligne [archive du ] [PDF]).
- (en) Douglas H. Erwin et Eric H. Davidson, « The last common bilaterian ancestor », Development, vol. 129, no 3, , p. 3021-3032 (PMID 12070079, lire en ligne).
- (en) Andrey Yu. Ivantsov, « Trace Fossils of Precambrian Metazoans "Vendobionta" and "Mollusks" », Stratigraphy and Geological Correlation, vol. 21, no 3, , p. 252–264 (DOI 10.1134/S0869593813030039, lire en ligne).
- (en) Andrey Yu. Ivantsov, « Paleontological evidence for the supposed precambrian occurrence of mollusks », Paleontological Journal, vol. 40, no 12, , p. 1552-1559 (DOI 10.1134/S0031030110120105, résumé).
- (en) N. J. Butterfield, « Hooking some stem-group "worms": fossil lophotrochozoans in the Burgess Shale », Bioessays, vol. 28, no 12, , p. 1161-1166 (PMID 17120226, DOI 10.1002/bies.20507)
- (en) N. J. Butterfield, « An Early Cambrian Radula », Journal of Paleontology, vol. 82, no 3, , p. 543-554 (DOI 10.1666/07-066).
Voir aussi
- (en) Référence Paleobiology Database : Kimberella Glaessner 1959