Halteria
Halteria est un genre de chromistes bactérivores et virovores[1] de la famille des Halteriidae.
| Domaine | Eukaryota |
|---|---|
| Règne | Chromista |
| Super-embr. | Alveolata |
| Embranchement | Ciliophora |
| Classe | Oligotrichea |
| Ordre | Halteriida |
| Famille | Halteriidae |
Espèces de rang inférieur
- H. grandinella
- H. bifurcate
- H. chorelligera
- H. cirrifera
- H. decemsulcata
- H. geleiana
- H. máxima
- H. mínima
- H. minuta
- H. oblonga
- H. ovata
- H. verrucosa
- H. viridis
- H. vorax
Les Halteria, parfois appelées « oligotriches sauteuses », sont des ciliés planctoniques communs dans de nombreux environnements d'eau douce. Les Halteria sont faciles à localiser en raison de leur abondance et de leur comportement. Les observations d'Halteria remonteraient au XVIIe siècle et à la découverte des microorganismes[2]. Au fil du temps, plus d'informations ont été établies sur leur morphologie et leur comportement, ce qui a conduit à faire évoluer leur classification.
Les espèces d'Halteria peuvent exister à la fois sous une forme trophique et une forme enkystée, mais sont le plus souvent décrites sous la forme trophique[3]. Elles peuvent être identifiées par leur mouvement de saut unique, activé par une rangée équatoriale de cirres rigides qui battent à l'unisson, permettant à l'organisme de se déplacer très rapidement vers l'arrière[4]. Les cellules d'Halteria sont à peu près en forme de dôme et, en plus des cirres équatoriaux, elles possèdent un collier de cils autour de l'ouverture buccale utilisée pour l'alimentation et la locomotion[4].
Les membres du genre Halteria sont hétérotrophes et servent d'importants bactérivores dans les habitats qu'ils occupent, où ils sont principalement la proie des métazoaires. Le rôle écologique important d'Halteria et sa stratégie de locomotion unique en font un genre d'intérêt dans différents domaines de la recherche en protistologie.
Étymologie
Le nom de la famille vient du genre type Halteria, dérivé du grec halter, saut, en référence au mode de déplacement par sauts de cet organisme.
Historique
Le genre Halteria est abondant dans de nombreux environnements d'eau douce[5]. L'ubiquité de ce genre est probablement la raison pour laquelle les observations remontent à des centaines d'années. La description originale du genre n'est pas clairement établie, mais il est possible que les observations d'Halteria remontent aux observations d'Antoni van Leeuwenhoek en 1675 en tant que quatrième animalcule observé dans un pot en terre rempli d'eau de pluie. L'organisme qu'il a observé était petit, rapide et immobile avant de changer rapidement de direction et de se déplacer tout droit, ce qui est cohérent avec le mouvement caractéristique d'Halteria[2].
Le nom Halteria est attribué à Félix Dujardin en 1840, qui a reclassé les espèces préalablement nommées Trichodina grandinella et Trichodina vorax respectivement par Müller et Ehrenberg, en H. grandinella et H. vorax[6]. Il a créé ce nouveau genre car ces deux espèces ne correspondaient pas à la sous-famille Vorticellina à laquelle le genre Trichodina appartenait[7]. Les descriptions d'Halteria à cette époque étaient encore assez vagues, se focalisant principalement sur le mouvement de saut rapide et la présence de cils buccaux[6].
En 1858, Édouard Claparède et Johannes Lachmann décrivent Halteria grandinella plus précisément, notant pour la première fois, que les cirres ne se trouvent que dans une ceinture équatoriale autour de la cellule. De nouveaux détails relatifs à la cavité buccale ont également été découverts ; Claparède et Lachmann ont observé qu'il y avait une échancrure dans une partie de l'appareil buccal et qu'à cet endroit aucun cil buccal n'est présent. Cela signifie que les cils buccaux forment un cercle incomplet autour de la cavité buccale et ne l'entourent pas complètement comme on le supposait auparavant[8].
Des questions sur la classification de Halteria se sont à nouveau posées ces dernières années. Ils ont été souvent classés comme membre du groupe des ciliés oligotrichs, car ils possèdent les cils oraux proéminents, disposés en un cercle incomplet, caractéristiques du groupe. Cependant, le séquençage récent du génome et l'analyse des ARN indiquent que Halteria serait plus étroitement lié aux Oxytrichidae qu'aux Oligotrichia, suggérant que la similitude de l'appareil oral avec les oligotrichs résulte d'une évolution convergente[9].
Description
Halteria peut exister sous forme trophique, ciliée, ou enkystée et la morphologie des cellules varie considérablement entre les stades[3].
Stade trophique
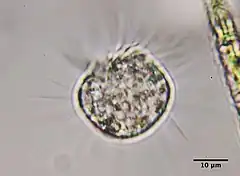
Au stade trophique, les cellules d'Halteria sont globulaires et mesurent entre 15 et 35 µm[3]. Les cellules possèdent à la fois des cils oraux et des cirres équatoriaux rigides[3]. Un collier de cils buccaux proéminents peut être trouvé à l'extrémité antérieure des cellules Halteria, entourant partiellement la cavité buccale. Cet appareil buccal se compose de quinze membranelles qui encerclent le péristome et de sept membranelles à l'intérieur de la cavité buccale[3].
Les cirres rigides de Halteria, parfois appelés poils sauteurs, mesurent chacun 15 à 25 µm de long[3]. Les cirres sont organisés de manière équatoriale autour des cellules en 7 à 10 rangées longitudinales[3]. Chaque rangée est à son tour organisée en quatre groupes de cirres. Lorsque des espèces de Halteria battent ces cirres à l'unisson, elles génèrent un mouvement de saut caractéristique suffisamment distinct pour Halteria que l'observation de ce mouvement a été considérée comme suffisante pour l'identification visuelle de Halteria[4].
Le cortex d'Halteria est composé de quatre membranes[3]. Deux de ces membranes, les membranes alvéolaires interne et externe, recouvrent les alvéoles plates qui se trouvent entièrement sous les deux membranes restantes[3]. La membrane cellulaire se trouve directement au-dessus de la membrane alvéolaire externe et recouvre toute la cellule, y compris les cils[3]. Le périlemme est la fragile membrane la plus externe qui ne recouvre que de petites parties de la cellule[3]. La fragilité du périlemme peut être à l'origine de cette répartition car il serait difficile à préserver[3]. Juste sous les membranes du cortex, la forme du corps d'Halteria est stabilisée par des microtubules dans une configuration en panier[3].
Dans les cellules d'Halteria, une vacuole contractile est située approximativement à mi-chemin entre les extrémités antérieure et postérieure de la cellule[3]. Les mitochondries de Halteria sont généralement sphériques avec des crêtes tubulaires[10]. Dans les mitochondries de H. geleiana, des microorganismes ont été observés dans la matrice[10]. Les microorganismes étaient en forme de bâtonnet et observés avec différentes longueurs et en différents nombres[10]. Aucune fonction ou origine n'est actuellement connue pour ces microorganismes ni s'ils sont parasites ou symbiotiques. Halteria a un micronoyau et un macronoyau avec de grands nucléoles en forme de bande[11] - [12]. Le macronoyau est de forme oblongue tandis que le micronoyau est plus globuleux[3].
Enkystement
Au fur et à mesure que les cellules d'Halteria passent du stade trophique au stade enkysté, leurs corps globulaires s'allongent initialement, principalement à l'extrémité antérieure, jusqu'à ce que la longueur de la cellule ait presque doublé[3]. En raison de l'allongement inégal, la cavité buccale est aplatie, les membranelles de l'appareil buccal se rapprochent du centre de la cellule et les rangées de cirres se rapprochent de l'extrémité postérieure de la cellule[3]. Pendant que la cellule s'étire, le cytoplasme développe des structures coniques de 5 µm de long[3]. Après cette étape d'allongement, les cellules s'arrondissent et une enveloppe muqueuse est extrudée. Également au cours de cette prochaine étape d'enkystement, les structures coniques formées dans le cytoplasme se fixent à la couche externe du kyste en développement, appelée ectocyste[3]. Une fois attachées à l'ectocyste, les structures coniques sont appelées lépidosomes[3]. Après l'enkystement, les kystes utilisent l'enveloppe muqueuse pour se fixer fermement à tout substrat disponible[3].
Écologie
Le genre Halteria se compose de ciliés d'eau douce qui vivent généralement un mode de vie planctonique. L'espèce Halteria grandinella est considérée comme cosmopolite, ce qui signifie qu'elle se trouve dans des habitats du monde entier[13]. D'autres espèces sont moins communes et donc moins bien définies, mais les descriptions fréquentes de Halteria grandinella donnent un aperçu du genre dans son ensemble. Les Halteria sont hétérotrophes et contrairement à de nombreux genres étroitement apparentés comme Pelagohalteria, ils n'ont pas d'endosymbiontes photosynthétiques. Halteria mange fréquemment des algues vertes qui, lorsqu'elles sont observées dans les vacuoles alimentaires, ont conduit à des erreurs de classification dans le passé lorsqu'elles ont été confondues avec des endosymbiontes[14].
Les espèces d'Halteria jouent un rôle particulièrement important dans de nombreux habitats d'eau douce en tant que bactérivores. Dans une étude qui a utilisé des bactéries marquées par fluorescence dans des étangs à poissons pour observer la bactérivorie du protistan, le pâturage des ciliés représentait 56 % du pâturage total du protistan et Halteria, ainsi que deux autres genres de ciliés, Pelagohalteria et Rimostrombidium, étaient responsables d'environ 71 % de la bactérivorie cilié total[15]. Halteria agit également comme une proie envers de nombreux animaux[16]. Il a été proposé que le comportement de saut caractéristique d''Halteria ait évolué comme une stratégie d'évasion pour éviter une telle prédation.
Les Halteria sont également virovores, capables de se se nourrir des virus présents[17] - [18].
Une grande partie de la recherche concernant Halteria se concentre sur leur mouvement et leur rôle écologique. Halteria est utilisé comme organisme modèle pour l'étude du mouvement de saut par battement ciliaire. On peut le trouver en abondance dans divers habitats d'eau douce interagissant avec d'autres organismes en tant que prédateurs u de proies[16] - [15].
Halteria passe la plupart du temps à l'arrêt ou à se déplacer doucement propulsée par les cils situés à leur extrémité antérieure[5]. Le mouvement de saut arrêté typique d'Halteria est le résultat d'un stimulus externe tel que des courants, ce qui a été mis en évidence en laboratoire[16]. Le comportement de saut chez Halteria mobilise 41 % des ressources du métabolisme de l'organisme[16], et donc y recourir trop fréquemment serait une utilisation dispendieuse de l'énergie.
Reproduction
Reproduction asexuée
Halteria peut se reproduire de manière asexuée par fission binaire transversale. Au cours de cette réplication, la plupart des cils des cellules filles est formée de novo[19]. La seule exception à cela est la ciliature orale de la cellule mère qui est héritée par la cellule fille proter[19]. Les cirres parentaux sont résorbés par la cellule pendant la division et les cirres des deux cellules filles sont produits de novo à partir d'anlagène cirral et la ciliature orale de la cellule fille opisthe est générée de novo par la formation d'un primordium oral à l'extrémité postérieure de la cellule[19]. Le macronoyau et le micronoyau se divisent au cours du processus, ce qui donne deux cellules filles génétiquement identiques à la cellule mère[19].
Reproduction sexuée
Les cellules Halteria peuvent se reproduire sexuellement grâce à un processus qui a été étudié spécifiquement chez H. grandinella[12]. Lors de la reproduction sexuée, les faces ventrales de deux cellules Halteria fusionnent. Divers changements de morphologie se produisent alors par divisions de maturation, notamment une diminution du nombre de cirres dans les deux cellules et la perte de membranelles buccales dans l'une des paires et la totalité de l'appareil buccal disparaît dans l'autre[12]. Les membranelles restantes sont partagées entre les cellules à l'extrémité antérieure[12]. Au niveau nucléaire, pendant la conjugaison, le fragment de macronoyaux d'origine et les micronoyaux mûrissent et se divisent trois fois, avec un seul dérivé des deux premières divisions continuant à se diviser, formant deux pronoyaux dans la troisième division[12]. Un pronoyau de chaque cellule est échangé et les deux qui se retrouvent dans chaque cellule fusionnent pour former le syncaryon[12]. Le synkaryon se divise deux fois avec un dérivé de chacune des deuxièmes divisions dégénérant et les dérivés restants devenant le nouveau micronoyau et le macronoyau analge. Une fois la division synkaryon terminée, les conjugués se séparent, générant deux cellules génétiquement distinctes des cellules mères et l'une de l'autre[12].
Liste des espèces
- H. grandinella
- H. bifurcate
- H. chorelligera
- H. cirrifera
- H. decemsulcata
- H. geleiana
- H. máxima
- H. mínima
- H. minuta
- H. oblonga
- H. ovata
- H. verrucosa
- H. viridis
- H. vorax
Liens externes
- (en) Référence BioLib : Halteria Dujardin, 1841 (consulté le )
- (en) Référence The Taxonomicon : Halteria Dujardin, 1841 (consulté le )
- (en) Référence World Register of Marine Species : taxon Halteria Dujardin, 1841 (+ liste espèces) (consulté le )
- Images
- (en) Halteria Grandinella sous le microscope (voir en ligne)
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Halteria » (voir la liste des auteurs).
- Antoine Gautherie, « Cet organisme ne mange que des virus et c’est une grande première », sur Journal du Geek, (consulté le ).
- Leeuwenhoek, A. V. (1677). Observations, communicated to the publisher by Mr. Antony van Leewenhoeck, in a dutch letter of the 9th Octob. 1676. here English'd: concerning little animals by him observed in rain-well-sea-and snow water; as also in water wherein pepper had lain infused. Philosophical Transactions of the Royal Society of London, 12(133), 821-831
- Foissner, W., Müller, H., & Agatha, S. (2007). A comparative fine structural and phylogenetic analysis of resting cysts in oligotrich and hypotrich Spirotrichea (Ciliophora). European Journal of Protistology, 43(4), 295-314
- Patterson, D.J. & Hedley, S., (1996). Freeliving and Freshwater Protozoa. CRC Press, Boca Raton, Florida, USA
- (en) J. H. Archbold et J. Berger, « A qualitative assessment of some metazoan predators of Halteria grandinella, a common freshwater ciliate », Hydrobiologia (en), vol. 126, no 2, , p. 97-102 (DOI 10.1007/BF00008675).
- F. Dujardin, Librairie Encyclopèdique de Roret, Paris, France, (OCLC 910488425, lire en ligne), « Histoire naturelle des zoophytes. Infusoires: comprenant la physiologie et la classification de ces animaux et la manière de les étudier à l'aide du microscope. »
- Pritchard A., « A History of Infusoria, Including the Desmidiaceae and Diatomaceae, British and Foreign: Enlarged and Revised by JT Anlidge, W », Whittaker and Company, vol. 27, no 54, , p. 445–446 (OCLC 969523285, PMCID 5182355, DOI 10.5962/bhl.title.101827, lire en ligne)
- R.É. Claparède, Études sur les infusoires et les rhizopodes, vol. 1, Geneva, Switzerland, Vaney, (OCLC 1253409690, lire en ligne)
- D.H. Lynn et M. Kolisko, « Molecules illuminate morphology: phylogenomics confirms convergent evolution among 'oligotrichous' ciliates », International Journal of Systematic and Evolutionary Microbiology, vol. 67, no 9, , p. 3676–82 (PMID 28829032, DOI 10.1099/ijsem.0.002060)
- Yamataka, S., & Hayashi, R. (1970). Electron microscopic studies on the mitochondria and intramitochondrial microorganisms of Halteria geleiana. Journal of Electron Microscopy, 19(1), 50-62.
- Petz, W., & Foissner, W. (1992). Morphology and Morphogenesis of Strobilidium caudatum (Fromentel), Meseres corlissi N. Sp., Halteria grandinella (Müller), and Strombidium rehwaldi N. Sp., and a Proposed Phylogenetic System for Oligotrich Ciliates (Protozoa, Ciliophora) 1. The Journal of Protozoology, 39(1), 159-176.
- Agatha, S., & Foissner, W. (2009). Conjugation in the spirotrich ciliate Halteria grandinella (Müller, 1773) Dujardin, 1841 (Protozoa, Ciliophora) and its phylogenetic implications. European Journal of Protistology, 45(1), 51-63.
- Foissner, W., Chao, A., & Katz, L. A. (2007). Diversity and geographic distribution of ciliates (Protista: Ciliophora). Protist diversity and geographical distribution (pp. 111-129). Springer, Dordrecht.
- Foissner, W. (1994). Progress in taxonomy of planktonic freshwater ciliates. Marine Microbial Food Webs, 8(1-2), 9-35
- Šimek, K., Jürgens, K., Nedoma, J., Comerma, M., & Armengol, J. (2000). Ecological role and bacterial grazing of Halteria spp.: small freshwater oligotrichs as dominant pelagic ciliate bacterivores. Aquatic Microbial Ecology, 22(1), 43-56. Wang, Chundi, et al. "Further analyses on the evolutionary “key‐protist” Halteria (Protista, Ciliophora) based on transcriptomic data." Zoologica Scripta 48.6 (2019): 813-825.
- Gilbert, J. J. (1994). Jumping behavior in the oligotrich ciliates Strobilidium velox and Halteria grandinella, and its significance as a defense against rotifer predators. Microbial Ecology, 27(2), 189-200.
- (en) John P. DeLong, James L. Van Etten, Zeina Al-Ameeli, Irina V. Agarkova et David D. Dunigan, « The consumption of viruses returns energy to food chains », Proceedings of the National Academy of Sciences, vol. 120, no 1, , e2215000120 (ISSN 0027-8424, DOI 10.1073/pnas.2215000120, lire en ligne)
- Michael Irving, « First "virovore" discovered: An organism that eats viruses », New Atlas, (lire en ligne [archive du ], consulté le )
- W. Song, « Studies on the cortical morphogenesis during cell division in Halteria grandinella (Muller, 1773) (Ciliophora, Oligotrichida) », Chinese Journal of Oceanology and Limnology, vol. 11, no 2, , p. 122–9 (DOI 10.1007/BF02850862, S2CID 84111747)