Dérive des invertébrés aquatiques
Dans le domaine de l’écologie des systèmes lotiques, la notion de dérive désigne le flux d’organismes (vivants ou morts[3]) emportés par le courant, et la « dérive des invertébrés aquatiques » désigne le cas particulier des migrations ou petits déplacements (actifs, c'est-à-dire comportementaux, ou passifs et accidentels) d’invertébrés dans le sens du courant[4].

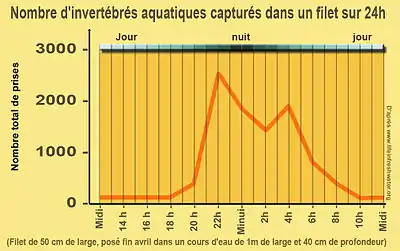



Elle concerne les invertébrés vivant sur le fond, dits benthiques, et les organismes planctoniques de pleine eau. Un phénomène assez similaire est constaté pour les alevins si des crues se produisent en leur présence. De nombreux poissons prédateurs se montrent également plus actifs aux heures auxquelles se produit cette dérive (aube, crépuscule).
Il s’agit parfois de l’une des formes de la migration animale et elle peut être associée à des phénomènes de migration verticale dans la colonne d'eau, souvent selon un cycle jour-nuit et/ou saisonnier.
Divers auteurs, dont Bournaud & Thibault en 1973, notent que « Il n'apparait jamais d'épuisement des zones amont par la dérive des organismes ». Selon eux, « les phénomènes compensateurs sont à rechercher surtout dans une production suffisante du benthos, mais aussi dans des déplacements actifs des organismes vers l'amont »[3]. Selon Bogadov (1984), grâce au flux de propagules et organismes adultes qui constituent la dérive, les communautés d’invertébrés et d’autres organismes benthiques sont plus stables et plus résilientes[5].
Le mot employé en allemand et en anglais pour désigner cette dérive est « drift », utilisé au moins depuis 1925 (par le naturaliste américain W.L. McAtee) dans ce contexte[6].
Espèces concernées
De très nombreuses espèces sont concernées ; dont en France concernant le benthos : des larves d'éphéméroptères (de la famille des Baetidae notamment), de plécoptères (Nemouridae notamment), de diptères (larves de simulies notamment, et parfois chironomes), des crustacés amphipodes tels que les aselles et gammares et des coléoptères aquatiques (larves et imagos de Dryopidae notamment). Les larves de trichoptères dérivent quand elles sont très jeunes, mais non pour celles qui construisent un fourreau minéral (une fois qu’elles l’ont construit) car ce fourreau est assez lourd pour les maintenir au fond. Le nombre d’hydracariens en dérive peut être sous-estimé si les mailles du filet sont trop grandes.
En période de crue, les filets utilisés par les biologistes pour évaluer cette dérive recueillent aussi de nombreux organismes ripicoles, semi-aquatiques ou non ripicoles et exogènes au cours d’eau (ex : 33 taxons qui représentaient 12,7 % de tous les organismes en dérive collectés dans le chenal principal de la Moyenne-Durance au milieu des années 1980.
Histoire scientifique
Ce thème a commencé à être scientifiquement étudié dans les années 1960-1970, dans différentes parties du monde et différents types de fleuves, rivières, torrents et ruisseaux, quand on a pris conscience de l’importance du flux de dérive d’invertébrés, de nuit notamment, et des enjeux pour les poissons carnivores ou omnivores et leurs alevins (salmonidés notamment[7]) ou encore d'enjeux écoépidémiologiques avec les mouvements d'organismes aquatiques ou semi-aquatiques vecteurs de maladies et zoonoses (Malaria par exemple).
En 1962, Waters confirme expérimentalement que l'accumulation d'invertébrés dans les filets de dérive résulte bien d'un déplacement permanent des organismes de l'amont vers l'aval, et qu'il ne s'agit pas du résultat d'une activité aléatoire et non-directionnelle des organismes situés dans le voisinage immédiat des filets[8].
En 1966, le même Waters a évalué la dérive à 0,001 à à, 5 g/m2/jour, ce qui correspondait dans ce cas à 1 centième à un tiers de toute la biomasse disparaissant chaque jour vers l'aval (suivant l'époque de l’année)[9]. Ce chiffre peut paraître élevé mais, dans ce même contexte, la production journalière était comprise entre 9,1 et 12,6 g/m2/j rien que pour Baetis vagans, et un turn-over important est connu pour de nombreux organismes[10], le dépeuplement induit par la dérive est donc dans une grande mesure compensé par les nouveaux individus amenés par la dérive elle-même et par les individus nés sur place[9]. Les conséquences de ces phénomènes ont suscité l'intérêt de divers scientifiques et justifié de nombreuses études, sans que le phénomène soit encore complètement cerné en termes d'enjeux écologiques notamment relatifs à la fragmentation écologique des cours d'eau et bassins versants, ou au contraire de liens artificiellement créés entre différents cours d'eau par les canaux, ou encore en termes de services écosystémiques.
Des informations sur la connectivité fonctionnelle et l’intégrité écologique du cours d’eau peuvent être obtenues en comparant les données de dévalaison d'alevins ou de poissons matures, la dérive du plancton et des macroinvertébrés benthiques. La comparaison peut aussi porter sur les dates et intensités de flux de matière organique, par exemple dans le cadre du «River Continuum Concept» (RCC).
Un travail fait sur le Rhône a conclu à un « fonctionnement contrôlé en priorité par l'amont, avec une relative indépendance des connectivités latérales, ses plaines d'inondation ne constituant pas apparemment une source de matières organiques à l'échelle de nos mesures ». Dans ce cas, les auteurs précisent que « les flux de dérive démontrent néanmoins, dans le contexte du « Patch Dynamics Concept » (PDC), le rôle fonctionnel des rives dans l'organisation des communautés et des populations de macro-invertébrés benthiques des écosystèmes lotiques de grands fleuves », ce qui invite à penser que les effets de l’artificialisation des berges de nombreux cours d’eau pourraient avoir été sous-estimés.
Caractérisation et mesure du phénomène
L'étude de ces dérives implique d'évaluer sur toute la largeur d'un cours d'eau (ou des cours d'eau d'un bassin versant) la densité et la masse des organismes (invertébrés dans le cadre du présent article) qui y passe et d'en expliquer la temporalité par deux types de facteurs :
- facteurs biotiques et endogènes (rythme nycthéméral, taxies, activité locomotrice, migrations, prédation, dérangement, phénomènes de compétition, etc.) ;
- facteurs abiotiques (température, teneur en oxygène, pollution éventuelle de l'eau).
Pour cela, les hydrobiologistes ont cherché à mieux caractériser son intensité, sa durée et sa périodicité, en mesurant notamment :
- « le taux de dérive », c'est-à-dire le nombre d’invertébrés dérivant par heure ou par jour[4] ;
- « l’intensité de dérive », qui est le nombre d'individus passant à travers 1 m2 de section en une heure[3]. Elle augmente généralement quand on s’approche du fond et dans les zones de fort courant, pouvant atteindre plusieurs millions d’individus par 24h sur une section de 10 à 20 m de large ;
- « la densité de dérive », qui s’évalue en comptant le nombre d’individus dérivants par unité de volume d’eau filtrée (en m3). Cette mesure permet des comparaisons entre différents sections du cours d’eau où le débit diffère[4] ;
- « la distance de dérive » ; son évaluation nécessite de marquer les individus et des techniques de capture/recapture, plus complexes et difficiles à mettre en œuvre dans les cours d’eau à fort courant ou en période de crue. C’est un paramètre encore mal connu, d’autant que certains organismes après s’être laissé dériver vont remonter vers l’amont (de 200 m par jour en migration de préémergence dans le cas de Leptophlebia cupida (éphéméroptère) étudié par Neave en 1930 ou de 30 à 65 cm par jour pour Gammarus bousfieldi étudié par Minckley en 1964. Les invertébrés peuvent aussi utiliser les contre-courants près des berges pour faciliter leur remontée vers l’amont Bournaud & Thibault (1973) [3] suggèrent d’ailleurs de ne pas omettre d’y installer des filets de contrôle quand on étudie les migrations des invertébrés aquatiques (les filets de dérive sont généralement placés là où le courant est le plus fort, c'est-à-dire loin des zones de contre-courant). Près des estuaires des espèces peuvent s’aider des courants de marées pour remonter vers l’amont après avoir dérivé[3] comme cela a été montré en automne pour Gammarus zaddachi (espèces univoltine) dans la Slack (petit fleuve côtier de la région Nord-Pas-de-Calais-Picardie). Hughes (1970) mesure (en laboratoire) des gammares qui remontent le courant à la vitesse de 22 mètres par heure ; cette remontée cessant au moment de la distribution de nourriture, et diminuant fortement quand le courant atteint 23 cm par seconde[2]. De grands crabes marins comme le « dormeur » utilisent une partie du courant de marée pour se déplacer contre le courant sur de grandes distances[11].
La mauvaise connaissance de cette distance de dérive pour la plupart des espèces marines ou de cours d'eau fait qu’on ne peut attribuer une origine aux organismes trouvés dans les filets ou filtres placés en travers du courant. - « Les heures et saisons de dérive » ; hormis pour quelques groupes qui migrent plus de jour, les pics de dérive pour la plupart des espèces sont constatés de nuit, et ils prennent deux formes que Bournaud & Thibault (1973) nomment « bigeminus » et « alternans » selon la forme de la courbe correspondant aux intensités de migration pour l'espèce[3].
Outils et méthodes de mesure
Ces mesures se font par différentes méthodes et en respectant certains principes nécessaires à la comparabilité :
- L'outil de base est le filet à maille fine dont il existe des modèles plus adaptés à certains courants ou pour l'échantillonnage au fond, en pleine eau ou encore en surface/subsurface. Mais Décamps et ses collègues ont en 1973 mis au point un échantillonneur de dérive associant un système Venturi et un manomètre différentiel permettant d’aussi connaitre le débit d'eau filtré[12].
- une méthode générale a été proposée par Elouard & Lévèque en 1975 [13]qui utilise trois filets à mailles fines disposés durant une certaine unité de temps en travers du courant, dans des zones de fort courant. Les individus et espèces piégées sont ensuite triés et comptés, permettant le calcul d’un IMD (« Indice moyen de dérive ») qui représente le nombre moyen théorique d'organismes dérivants, par seconde et par mètre cube d'eau, pour le cours d'eau étudié. Pour bien prendre en compte la dérive de surface, le filet doit dépasser le niveau de l’eau. Pour ne mesurer que la dérive de fond, il est plaqué au fond. le filet peut aussi être placé sous une chute d’eau (et éventuellement alors mesurer la totalité de la dérive sans que le débit soit modifié par la présence du filet dont les mailles sont souvent très fines (cas du filet à plancton).
- Bournaud & Thibault signalent dans leur synthèse bibliographique de 1973 plusieurs auteurs : Müller (1965) [14], Küreck (1967) [15], et Bishop (1969) qui ont ajouté à leurs protocoles expérimentaux un diispositif permettant de récupérer le contenu du filet par tranches horaires (1 à 3h selon les cas) ce qui permet de mesurer les variations horaires sur un même point [3]. Ils recommandent ce type de mesure séquencée qui seule permet d'évaluer l'importance des variations nycthémérales (pouvant d'ailleurs être perturbés par un clair de lune).
- Pour être comparables d'un cours d'eau à l'autre ou d'une partie d'un cours d'eau à l'autre, le filet ne doit pas se colmater durant la période de mesure ce qui implique la présence d’un opérateur surveillant et nettoyant le filet si l’eau est chargée de débris, feuilles, algues, etc. De plus, les durées et heures de poses des filets doivent être identiques.
- L'heure de pose du filet a une grande importance, la dérive pouvant être plus de 10 fois plus importante après le coucher du soleil qu'à midi.
- La saison a une importance qui varie selon la zone climatique considérée (d’autant plus qu’on s’éloigne de l’équateur et qu’on se rapproche des pôles ou qu’on monte en altitude). La survenue de crues en amont, avant ou durant le piégeage, doit également être prise en compte.
- Une méthode de mesure des effets d'un toxique sur la dérive été formalisée par Dejoux en 1975, puis précisée par Dejoux et Trouvat en 1976 ; Elle consiste à utiliser des gouttières semi immergées dans laquelle sont disposés des sédiments du cours d'eau et leurs organismes. Cette gouttière est ensuite exposée au passage du toxique dans l'eau courante et on mesure à sa sortie la quantité d'organismes qui se sont décrochés et sont emportés à la dérive.
- Les comparaisons de biomasse impliquent de bien préciser comment la masse des organismes a été mesurée. En général il s’agit du poids sec ; après 48 h de séchage à 38 °C (pour Bailey en 1966 [16]) ou 70 °C (pour Anderson et Lehmkuhl en 1968 [17]) ou après une légère centrifugation pour Waters en 1966[9].
Compensation constante de la dérive
S’ils n’étaient pas naturellement compensés, des phénomènes importants de dérive (fréquemment constatés) tendraient à vider l’amont et certains compartiments des cours d’eau de leurs invertébrés.
La plupart des cours d’eau sont en fait régulièrement ou constamment « repeuplés » par des propagules venus de l’amont (ou de l’aval, remontés par des poissons, oiseaux ou mammifères par exemple, on parle alors de zoochorie de l’aval vers l’amont).
Ainsi la dérive d’un grand cours d’eau est compensée par la dérive d’invertébrés provenant de l’amont, mais aussi et très significativement provenant de bras annexes, de chenaux latéraux et de zones périodiquement inondées.
Selon le naturaliste Karl Müller[18] - [19] la recolonisation de l'amont pourrait se faire par des imagos qui remontent vers la source en volant puis pondent plus en amont. Bournaud & Thibault (1973) rappellent aussi que chez certaines espèces (gammares), il a aussi été constaté que les femelles ovigères se laissaient moins dériver que les mâles et même qu'elles remontent vers l'amont.
Ce phénomène explique en partie la résilience écologique de cours d’eau curés, momentanément asséchés ou momentanément gravement pollués. Il est aussi constaté dans des canaux ou cours d’eau semi-artificialisés ou aménagés avec de grands barrages (comme la Moyenne-Durance par exemple[4]). Il est quantitativement parfois très important : ainsi Müller a en 1954 calculé que 4,550 kg d'invertébrés avaient en seulement 11 jours du mois de juin recolonisés 150 m de ruisseaux auparavant entièrement nettoyés. Cette recolonisation ne peut provenir que d'organismes apportés par le courant à partir de l'amont, ou provenant de l'aval à la suite d'une remontée du cours d'eau vers sa source.
Ce qui contrôle la dérive des invertébrés
De nombreuses observations in situ ont montré que hors évènements accidentels, les organismes se laissent volontairement dériver en rejoignant eux-mêmes la zone de plus fort courant et en y retournant s’ils sen éloignent ou après une courte période.
La dérive d’organismes benthiques est néanmoins sous le contrôle de plusieurs phénomènes souvent conjoints ou successifs.
- la microrépartition des invertébrés. La microdistribution des espèces et individus varie selon la saison, l'altitude, et divers facteurs abiotiques, etc.[20] - [21]
- le rythme nycthéméral. Le nycthémère influence naturellement et de manière majeure le comportement de la plupart des organismes, avec toutefois des pics horaires de densité d’individus en dérive (taux de dérive par exemple multiplié par 10 dans les 3 ou 4 heures suivant le coucher du soleil). L’heure du pic nocturne change légèrement à significativement selon les espèces considérées, mais aussi selon les sections et la naturalité du cours d’eau.
La très grande majorité des invertébrés ne se laissent dériver que de nuit (sauf dérive accidentelle) : la dérive est par exemple environ 80 fois plus intense de nuit chez Baetis rhodani et dix fois chez plusieurs espèces de plécoptères [3].
Chez la plupart des espèces le pic de dérive suit le coucher du soleil. Il existe quelques exceptions avec par exemple en zone tempérée : des ostracodes, des oligochètes, des Limnephilides (Oligophlebodes sigma et Psychoglypha alaskensis), un Brachycentridé (Brachycentrus americanus), des larves et imagos de coléoptères Parnidés et quelques larves de Chironomidés et simulies [3]. La lumière ambiante étant souvent un paramètre déterminant[22], les éclairages artificiels situés sur les ponts et en bordure de cours d'eau pourraient peut-être perturber la dérive
- rythme lunaire. Après l’hypothèse posée par Waters en 1962, son compatriote Anderson[23] a clairement confirmé en 1966 que la lumière de la Lune (en phase pleine lune notamment) inhibe fortement la dérive de certaines espèces[24]. Il s'agit sans doute d'un comportement favorisé par la sélection naturelle car il expose a priori fortement les organismes à une prédation nocturne accrue. En 1960, Beeton avait déjà étudié un phénomène similaire concernant les migrations verticales de la crevette d'eau douce Mysis relicta[25], et un phénomène similaire est également démontré pour des coléoptères aquatiques non concerné par la dérive car vivant dans les mares.
- Saisonnalité. L'heure du « maximum de dérive » peut changer avec les changements de saisons[3]. Ce pic est par exemple pour certaines espèces observé en début de nuit de décembre à avril, mais pour la même espèce se fera en fin de nuit au mois de juin selon Elliott et Minshall (1968)[26].
- Crues/décrues. Les crues suscitent une dérive locale, mais aussi une « dérive exogène », c'est-à-dire constituée d’organismes provenant du proche bassin-versant et non plus seulement de l’amont du cours d'eau. Ce phénomène a par exemple été étudié dans le Massif central par Neveu et Echaubard (1975)[27] ;
- stress anoxique. Une désoxygénation expérimentale partielle d'un ruisseau des Pyrénées centrales a permis en 1975 de démontrer l’existence d’un seuil d’oxygénation de l'eau (environ 4 ppm/L dans le contexte de cette expérience). Sous ce seuil la dérive des invertébrés benthiques s'accroît significativement (avec toutefois des différences par espèce et selon « l'intensité et la durée de la désoxygénation »[28]. Selon Lavandier & Capblancq, les auteurs de cette expérience « les variations quotidiennes des teneurs en 02 dissous que présentent naturellement les eaux courantes, ne paraissent entraîner aucune réaction de la faune si elles se situent au-dessus de ce seuil »[28] ;
- Stress thermique. Une température inhabituelle ment chaude de l'eau augmente la dérive, mais avec des seuils de sensibilité variant selon les espèces et leur stade de développement. Cet effet a par exemple étudié pour le gammare Gammarus pseudolimnaeus Bousfield et pour Baetis vagans par Wojtalik & Waters (1970)[29] ; Les quelques espèces qui dérivent principalement de jour semblent le faire notamment sous le contrôle de la température de l’eau[30] ;
- stress chimique ; Une pollution organique[31] - [32] ou acide[33] peuvent déclencher une augmentation de la dérive ;
- stress écotoxique ; À titre d'exemple, l'application de pesticides de démoustication en zone humide aux abords de l’eau ou dans l’eau affecte aussi des invertébrés non-ciblés[34] - [35]. Ces derniers peuvent rapidement mourir et être emportés, ou sous l'effet du stress peuvent se décrocher de leur substrat et se laisser emporter par le courant. Elles sont alors parfois massivement retrouvées en aval dans les filets de mesure de la dérive d’invertébrés. Ainsi lors d’une étude faite en Afrique de l'Ouest, Dejous et Elourard ont montré en 1977 qu’après un premier traitement aquatique à l’Abate (anti-moustique et anti-larves de simulies), jusqu’à 50 % des invertébrés du milieu sont retrouvés morts (rapidement après l’application)[36], mais que si les traitements se répètent, une sélection d’espèces ou d’individus résistants se fait et qu’on observe alors moins de décrochements et d’effets sur la dérive ;
- « Effet surpopulation » ? . C'est une hypothèse qui est discutée : Selon Dimond (1967) le phénomène de dérive n'apparaitrait qu'à partir d'un certain seuil de population benthique ou dépassement de la capacité écologique d'accueil du milieu, et elle serait donc un moyen pour les espèces de localement limiter leur surpopulation[37], et Waters[9]. Il se base pour fonder cette hypothèse sur le fait que dans certains ruisseaux traités par des insecticides, la dérive n'est revenue à la normale que trois ans après la réapparition du benthos[9]. Au contraire et peu après, Eliott et Minshall (1968) ont estimé qu'en présence de phénomènes de dérive on ne peut généralement pas parler de dépassement de la capacité biotique, car (hormis pour la partie située très en amont des cours d'eau) la dérive correspond simplement à une translation d'une partie de la biomasse, et non à sa disparition de la section concernée du cours d'eau.
De plus d'autres études montreront ensuite qu’une partie des individus qui se sont laissés dérivés sont retrouvés en amont après avoir remonté le courant, parfois sur de grandes distances (phénomène également constaté en mer : des crabes dormeurs (Cancer pagurus) marqués ont ainsi été retrouvés jusqu’à plus de 260 km de leur point de capture) et très loin de leur premier lieu de capture (en amont par rapport à la dérive résiduelle du courant de marée[11] ; une explication plausible est que la femelle doit « compenser, en l'anticipant, la dérive que subissent les larves au cours de leur vie pélagique ».
Cas particuliers
- Les dérives catastrophiques.. Ce sont des dérives en quelque sorte « accidentelles ». Elles ont d’abord été signalées par Waters dans les années 1960[38] - [8] puis par beaucoup d’autres auteurs. Waters cite comme exemple une forte croissance du nombre d'invertébrés arrivant dans ses filets (décuplement du nombre de larves de simulies) après que des enfants aient piétiné le fond de la rivière plus en amont. De même après la débâcle des glaces ou même après une pluie assez forte pour engendrer une crue, phénomène observé par de nombreux auteurs et décrit au moins depuis 1928 par Needham [39].
- Les cours d'eau temporaires. Ils connaissent aussi des phénomènes de dérives, mais conditionnés par la présence périodique de l'eau, et la force du courant quand il est présent. La dérive contribue elle-même à la recolonisation plus ou moins rapide du lit après le retour de l'eau, permettant le retour d'un fonctionnement s'apparentant - dans une certaine mesure - à ceux des cours d'eau permanents[40]
- Les systèmes karstiques. Dans ces cas, la dérive se fait moins linéaire et s'enfonce sous la terre, souvent pour ne jamais réapparaître dans les résurgences. Ce type de situation a notamment été étudié par Décamp (1973) [41] - [42]dans les karsts du Baget (Pyrénées-Atlantiques). Une partie de la dérive disparait dans le milieu souterrain sans être retrouvée au niveau des résurgences[43] ; En période de crues plus de 90 % des invertébrés entrant dans les karsts y « restent piégé ». Le "pic" (en nombre d'individus et biomasse) se produit durant les premiers jours de crue et il correspond au pic de « matière organique globale transportée par litre d'eau » avec une fraction de particules de grandes et moyennes taille particulièrement bien représentée, alors qu'au contraire en sortie du réseau karstique c'est la matière organique dissoute qui constitue la fraction la plus abondante[43]. Dans ce cas, lors d'une crue hivernale (), le nombre d'invertébrés en dérive a varié de 1 000 à 1 800 individus/heure soit, pour les 8 heures d'échantillonnage : 168 000 invertébré, sachant que le filet ne filtrait que 1/15 environ du débit total du ruisseau entrant dans le karst)[43].
Ces "pertes" sont l'un des éléments à prendre en compte lors de l'étude du cas particulier des écotones eau de surface/eaux souterraines quand ils concernent des zones karstiques[44].
- Les cours d'eau aménagés : Les canaux, les aménagements hydroélectriques, les barrages de retenues d'eau et leurs éclusées interfèrent avec les phénomènes de dérive[45] - [46]. Ceci se produit également, et conjointement souvent pour les alevins de poissons ( salmonidés par exemple)[47].
Une étude s'est intéressée à l'influence sur la dérive des copépodes de la configuration d'un cours d'eau canalisée et de son affluent plus naturel ; dans ce cas les échantillonnage ont montré que les copépodes constituaient 0,75 % de la dérive dans le cours principal de la Moyenne-Durance vers 1985, mais plus de 10 fois plus (10,7 %) dans son chenal secondaire (lequel est relié à un petit marais). Parmi les copépodes, une espèces (Macrocyclops albidus) était largement dominante, constituant 88 % des individus en dérive, devant Acanthocyclops robustus (10 %) et d'autres espèces moins représentées (Eucyclops serrulatus...).
L'effet des barrages hydroélectriques sur la dérive des invertébrés a par exemple été bien étudiés pour leurs effets dans la Haute Neste d'Aure[48] - [49].
Phénomènes homologues dans d'autres groupes taxonomiques
Ils sont observés chez les vertébrés que sont les poissons, mais aussi, avec des rythmes différents chez les algues aquatiques et certaines plantes aquatiques. On parle par exemple de dérive algale[50]. On retrouve dans l’analyse du phénomène des paramètres pouvant évoquer la dérive des invertébrés (influence de perturbations naturelles (température, luminosité, pression de broutage), stress anthropiques, nature du substrat, variations nycthémérales, stabilité du milieu, biomasse en place…). La dérive de propagules d’algues épilithique contribue à la recolonisation du milieu après une crue vive ou un long assec.
Certaines propagules de plantes aquatiques font aussi l’objet d’une dérive jouant un rôle important pour leur dispersion.
En mer la dérive de grands laminaires décrochés du fond contribue à diffuser les invertébrés qui leur sont associés, par exemple entre l'Afrique du Sud et Sainte-Hélène [51].
Notes et références
- William D. Pearson & Donald R. Franklin (1968), Some Factors Affecting Drift Rates of Baetis and Simuliidae in a Large River Ecology, Vol.49, no 1 (janvier 1968), p. 75-81 ; Ed : Ecological Society of America ; DOI:10.2307/1933562 7 pages (résumé et 1re page
- Hugues D.A (1970) Some factors affecting drift and upstream movements of gammarus pulex ; Ecology, 51, 2, 301-305
- Bournaud, M., & Thibault, M. (1973) La dérive des organismes dans les eaux courantes (étude bibliographique pour la littérature publiée avant 1970) ; Ann. Hydrobiol., 1973, 4(1), II-49 (1).
- Prevot G & Prevot R (1986), Impact d'une crue sur la communauté d'invertébrés de la Moyenne Durance. Rôle de la dérive dans la reconstitution du peuplement du chenal principal, In Annales de limnologie (Vol. 22, no 1, p. 89-98). EDP Sciences.
- Bogatov V (1984). "Influence du flux de benthos dans les processus de productivité biologique dans les rivièresInfluence of benthos flux in the process of stream biological productivity."(Notice/résumé Inist-CNRS, Èkologiâ no3, pp. 52-60 (18 ref.) ISSN 0367-0597
- Mc Atee W.L (1925) Notes on drift, begetable balls, and aquatic insectes as a food product of insland waters. Ecology, 6, 3, 288-302
- Waters, T. F. (1969). Invertebrate drift-ecology and significance to stream fishes. In Symposium on salmon and trout in streams (p. 121-134). Institute of Fisheries, University of British Columbia.
- Waters T.F (1962) Interpretation of invertebrates drift in streams. Ecology, 46, 3, 327-334 (https://www.jstor.org/stable/1936336 résumé])
- Waters T.F (1966) Production rate, population density and drift of a stream invertebrate Ecology, 47, 595-605
- Waters T.F (1969), The turnover ratio in production ecology of freshwater invertebrates. American Naturalist, 173-185.
- Contribution à l'étude de la biologie du crabe-tourteau Cancer pagurus sur les côtes de Bretagne sud ; Anne Le Foll, Ifremer
- Décamps, H., H. Laville, and D. Trivellato. "Utilisation d'un venturi pour un échantillonneur de dérive en eau courante." Annales de Limnologie. Vol. 9. No. 02. EDP Sciences, 1973. (résumé)
- Elouard JM & Lévèque C (1975) Observations préliminaires sur la dérive des invertébrés et des poissons dans quelques rivières de cöte d'Ivoire, ORSTOM-OMS, n° 394, 14 p
- Müller (1965), An automatic stream drift sampler, Limnol. Oceanogr, 10, 483-485
- Küreck A (1967) Uber die tagesperiodische Ausdrift von Niphargus aquilex schellenbergi KARAMAN, aus Quellen. Z. Morphol. ökol. Tiere, 58, 3, 247-262
- Bailey RG (1966) Observations on the nature and importance of organic drift in a Devon river. Hydrobiologia, 27, 3/4, 353-367
- Anderson NH & lehmkuhl DM (1968) Catastropic drift of insectes in a woodland stream. Ecology, 49, 2, 198-206
- Müller Karl (1954), Investigations on the organic drift in North swedish Streams. Rep. inst. Freshwat Res. Drottningholm, 34, 133-148
- Müller Karl (1966), Die Tagesperiodik von Fliesswasserorganismen. Z. Morphol. ökol. Tiere, 56, 1, 93-142
- Lavandier P & Dumas J (1971), Microrépartition de quelques espèces d'invertébrés benthiques dans des ruisseaux des Pyrénées centrales. Annls. Limnol., 7 (1) ; 7-23.
- Décamps, H., G. Larrouy, and D. Trivellato. (1975) "Approche hydrodynamique de la microdistribution d'invertébrés benthiques en eau courante." Annales de Limnologie. Vol. 11. No. 01. EDP Sciences (résumé)
- Holt C.S & Waters T.F (1967) Effect of light intensity on the drift of stream invertebrates. Ecology, 225-234.
- Department of Entomology, Oregon State University, Corvallis, Oregon
- Anderson N.H (1966) « Depressant effect of moonlight on activity of aquatic insects » ; Letters to Nature, Nature 209, 319 - 320 (15 janvier 1966) ; Doi:10.1038/209319a0 (résumé)
- Beeton, A. M. , J. Fish. Res. Bd. Canada, 17, 517 (1960)
- Eliott JM & Minshall GM (1968), The invertebrate drift in the river Duddon, English Lake District, Oikos, 19, I, 39-52
- Neveu (A.) & Echaubard (M.) (1975) « La dérive estivale des invertébrés aquatiques et terrestres dans un ruisseau du Massif Central » ; la Couze-Pavin. Ann. Hydrobiol., 6 : 1-26.
- Lavandier P & Capblancq J. (1975), Influence des variations d'oxygène dissous sur les invertébrés benthiques d'un ruisseau des Pyrénées centrales; Annls Limnol. 11 (1) : 101-106
- Wojtalik T.A & Waters T.F (1970) Some effects of heated water on the drift of two species of stream invertebrates. Transactions of the American Fisheries Society, 99(4), 782-788
- Waters T.F (1968) Diurnal periodicity in the drift of a day-active stream invertebrate. Ecology, 49(1), 152-153 (résumé).
- Pegaz-Maucet, Dominique (1980) Impact d'une perturbation d'origine organique sur la dérive des macro-invertebres d'un cours d'eau : comparaison avec le benthos. Thèse de doctorat.
- Giudicelli J et al. (1986), "Impact d'une perturbation polluante d'origine organique sur les populations et les communautés d'invertébrés benthiques et en dérive d'une rivière méditerranéenne, le Tavignano (Corse)." Ecologia mediterranea 12.1-2: 31-63
- Allard, M., and G. Moreau. "Influence d’une acidification expérimentale en milieu lotique sur la dérive des invertébrés benthiques." Verh. Internat. Verein. Lol 22 (1984): 1793-1800.
- Dejoux C (1982), "Recherche sur le devenir invertébrés dérivant dans un cours d'eau tropical à la suite traitements antisimulidiens au téméphos." Rev. Francais Sci. de l'eau 1 : 267-283.
- Gibon, François-Marie, and Jean-Jacques Troubat. "Effets d'un traitement au chlorphoxim sur la derive des invertebres benthiques." (1980).
- Dejous C & Elouard J.M (1977) Action de l'abate sur les invertébrés aquatiques ; cinétique de décrochement à court et moyen terme, Cahiers de l’O.R.S.T.O.M., série. Hydrobiologie, Vol. XI, no 3, 1977: 217-230. (disponible sur le site de l’IRD)
- Dimond J.B (1967) “Evidence that drift of strem benthos is density related”. Ecology, 48, 5, 855-857
- Waters T.F (1962) Diurnal periodicity on the drift of stream invertebrates ; Ecology, 43, 2, 316-320 (Lien vers la Revue, sans résumé).
- Needham P.R (1928) A quantitative study of the fish food supply in selected areas. Suppl. 17th Ann. Rept. N.Y. Cons Comm, 192-284
- Legier P (1984) "Importance de la dérive dans la mise en place des communautés d'invertébrés dans les cours d'eau temporaires de la Provence calcaire." Ecologia mediterranea 10.1-2 : 3-44 (http://cat.inist.fr/?aModele=afficheN&cpsidt=8899842 Notice/résumé Inist-CNRS]).
- Décamps H & Rouch R (1973), Le Système karstique du Baget : Premières estimations sur la dérive des invertébrés aquatiques d'origine épigée. CNRS
- Rouch R (1979) "Le système karstique du Baget. VIII. La communauté des Harpacticides. Les apports au sein du système par dérive catastrophique." Annales de Limnologie. Vol. 15. No. 03. EDP Sciences (résumé)
- Décamps H & Laville H (1975) "Invertébrés et matières organiques entraînés lors des crues à l'entrée et à la sortie du système karstique du Baget" ; Annales de Limnologie. Vol. 11. N°3. EDP Sciences résumé)
- Vervier Ph & J. Gibert (1991) "Dynamics of surface water/groundwater ecotones in a karstic aquifer." Freshwater Biology 26.2 (1991): 241-250. résumé
- Céréghino Régis (1997), Influence des éclusées hydroélectriques sur la structure et la dynamique des populations d'invertébrés d'une rivière pyrénéenne de moyenne montagne. Diss. (http://cat.inist.fr/?aModele=afficheN&cpsidt=180889 résumé])
- Lauters, François (1995), Impacts sur l'écosystème aquatique de la gestion par éclusées des ouvrages hydroélectriques. Étude de quelques cours d'eau et analyse des phénomènes mis en jeu. Diss. (résumé)
- Liebig H, Lim P & Belaud A (1998), "Influence du débit de base et de la durée des éclusées sur la dérive d'alevins de truite commune: expérimentations en canal semi-naturel." Bulletin Français de la Pêche et de la Pisciculture 350-351: 337-347.
- Gazagnes Gérard (1983) Contribution à l'étude de la dérive des invertébrés sur la haute Neste d'Aure; impact des aménagements hydro-électriques. Thèse de 3e cycle en Hydrobiologie, soutenue à l'Université de Toulouse 3
- Holmière, Dominique (1988), La dérive des invertébrés sur la moyenne et basse Neste d'Aure (Hautes Pyrénées) : impact des aménagements hydroelectriques. Thèse de doctorat, soutenu à l'université de Toulouse 3 (lien vers notice www.theses.fr)
- Rolland, Thierry (1995). Adaptation des méthodes d'échantillonnage et d'analyse en rivières méditerranéennes du Sud-est de la France: étude de l'hétérogénéité spatio-temporelle de l'épilithon et de la dérive algale. Thèse de doctorat, (Notice-résumé Inist-CNRS) . Université d'Aix-Marseille 3, Aix-en-Provence
- Arnaud, Françoise & al. "Transport d'invertébrés benthiques entre l'Afrique du Sud et Sainte-Hélène par les laminaires (Phaeophyceae)." Bulletin du Museum National d’Histoire Naturelle Paris Séries 3.384 (1976): 49-55.
Voir aussi
Articles connexes
Bibliographie
- Allard, M., and G. Moreau. "Influence d’une acidification expérimentale en milieu lotique sur la dérive des invertébrés benthiques." Verh. Internat. Verein. Lol 22 (1984): 1793-1800.
- Bournaud M & Thibault M (1973) La dérive des organismes dans les eaux courantes ; Ann. Hydrobiol., 1973, 4(1), II-49 (1).
- Gibon, François-Marie & Troubat, Jean-Jacques 1980), "Effets d'un traitement au chlorphoxim sur la dérive des invertébrés benthiques"
- Hall R.J, Waters T.F & Cook E.F (1980) The role of drift dispersal in production ecology of a stream mayfly. Ecology, 37-43 (résumé).
- Müller, Karl (1974) Stream drift as a chronobiological phenomenon in running water ecosystems. Annual Review of Ecology and Systematics, 309-323 (lien vers la 1re page).
- Pegaz-Maucet, Dominique (1980). Impact d'une perturbation d'origine organique sur la derive des macro-invertebres d'un cours d'eau: comparaison avec le benthos. Thèse de doctorat.
- Rader, Russell B (1997), "A functional classification of the drift: traits that influence invertebrate availability to salmonids." Canadian Journal of Fisheries and Aquatic Sciences 54.6 : 1211-1234.
- Troubat Jean-Jacques et al. (1982) "Effets d'un épandage sur le cycle de dérive et les densités d'insectes benthiques." (IRD).
- Vignes J-C. "Rythmes alimentaires de jeunes saumons atlantiques (Salmo salar L.) et relations trophiques avec la dérive des invertébrés." Vie et milieu 49.4 (1999): 293-300.
- Waters T.F (1966), Production rate, population density, and drift of a stream invertebrate. Ecology, 595-604 (résumé)
- Waters T.F (1968). Diurnal periodicity in the drift of a day-active stream invertebrate. Ecology, 49(1), 152-153 (résumé).
- Waters T.F (1969) Diel patterns of aquatic invertebrate drift in streams of northern Utah. Proc. Utah Acad. Sci. Arts Lett, 46(pt 2), 109-130.
- Waters T.F (1972) The drift of stream insects. Annual review of entomology, 17(1), 253-272 (Lien vers 1re page).
- William D. Pearson and Donald R. Franklin (), Some Factors Affecting Drift Rates of Baetis and Simuliidae in a Large River ; Ecology, Vol.49, no 1 (), p. 75-81 ; Ed : Ecological Society of America ; DOI:10.2307/1933562 7 pages (résumé et 1re page)