Déplacement de caractères
Le déplacement de caractères est le phénomène dans lequel les différences entre des espèces similaires dont les distributions se chevauchent géographiquement sont accentuées dans les régions où les espèces coexistent, mais sont minimisées ou perdues lorsque les distributions des espèces ne se chevauchent pas. Ce schéma résulte d'un changement évolutif induit par la compétition biologique entre les espèces pour une ressource limitée (par exemple la nourriture). La raison d'être du déplacement de caractères découle du principe d'exclusion compétitive, également appelé loi de Gause, qui soutient que pour coexister dans un environnement stable, deux espèces concurrentes doivent avoir une niche écologique différente ; sans cette différenciation, une espèce éliminera ou exclura l'autre par la compétition.
Le déplacement de caractères a été expliqué explicitement pour la première fois par William Brown Jr. et E. O. Wilson en 1956[1] : « Deux espèces étroitement apparentées ont des aires de répartition qui se chevauchent. Dans les zones des aires de répartition où une espèce est présente seule, les populations de cette espèce sont similaires aux autres espèces et peuvent même être très difficiles à distinguer d'elle. Dans la zone de chevauchement, où les deux espèces se rencontrent, les populations sont plus divergentes et plus faciles à distinguer, c'est-à-dire qu'elles "s'éloignent" l'une de l'autre relativement à un ou plusieurs caractères (d'où la notion de "déplacement"). Les caractères impliqués peuvent être morphologiques, écologiques, comportementaux ou physiologiques ; ils sont supposés être basés sur la génétique. »
Brown et Wilson ont utilisé le terme déplacement de caractères pour désigner à la fois du déplacement de caractères reproductifs, ou renforcement des barrières reproductives, et du déplacement de caractères écologiques qu'entraîne la compétition[1]. Comme le terme "déplacement de caractères" est couramment utilisé, il fait généralement référence aux différences morphologiques dues à la compétition. Brown et Wilson considéraient le déplacement de caractères comme un phénomène impliqué dans la spéciation, déclarant : « Nous pensons qu'il s'agit d'un aspect commun de la spéciation géographique, apparaissant le plus souvent comme un produit de l'interaction génétique et écologique de deux (ou plus) espèces apparentées nouvellement apparues dans l'évolution [dérivées de la même espèce parentale immédiate] pendant leur période de premier contact. » Bien que le déplacement de caractères soit important dans divers scénarios de spéciation[2], comprenant par exemple les radiations adaptatives rencontrées chez les faunes de poissons cichlidés dans les lacs de rift d'Afrique de l'Est[3], il joue également un rôle important dans la structuration des communautés. Il joue également un rôle dans la spéciation par le renforcement de telle sorte que les populations allopatriques qui se chevauchent en sympatrie présentent une plus grande divergence des traits[4]. Les résultats de nombreuses études apportent la preuve que le déplacement de caractères influence souvent l'évolution de l'acquisition des ressources chez les membres d'une guilde écologique[5].
La libération compétitive, définie comme l'expansion d'une niche écologique en l'absence d'un concurrent, est essentiellement l'image miroir du déplacement des caractères[6]. Brown et Wilson l'ont également décrit[1] : « Deux espèces étroitement apparentées sont distinctes là où elles se trouvent ensemble, mais lorsqu'un membre de la paire se trouve seul, il converge vers le second, même au point d'être presque identique à lui dans certains caractères. »
Développement conceptuel
« Le déplacement de caractères est la situation dans laquelle, lorsque deux espèces d'animaux se chevauchent géographiquement, les différences entre elles sont accentuées dans la zone de sympatrie et affaiblies ou complètement perdues dans les parties de leur aire de répartition situées en dehors de cette zone. »[1] Alors que le terme "déplacement des caractéristiques écologiques" est apparu pour la première fois dans la littérature scientifique en 1956, l'idée a des racines antérieures. Par exemple, Joseph Grinnell, dans l'article classique qui exposait le concept de niche écologique, a déclaré : « Il est bien sûr axiomatique que deux espèces régulièrement établies dans une même faune n'ont pas exactement les mêmes exigences en termes de niche. »[6] L'existence du déplacement de caractères prouve que les deux espèces ne se chevauchent pas complètement dans leur exigence de niche.
Après la diffusion du concept, le déplacement de caractères a été considéré comme un moteur important dans la structuration des communautés écologiques, et les biologistes en ont identifié de nombreux exemples. À la fin des années 1970 et au début des années 1980, cependant, le rôle de la compétition et du déplacement de caractères dans la structuration des communautés a été remis en question et son importance considérablement réduite[7]. Beaucoup ont trouvé les premiers exemples peu convaincants et ont suggéré qu'il s'agissait d'un phénomène rare. Les critiques des études antérieures comprenaient le manque de rigueur dans les analyses statistiques et l'utilisation de caractères mal rationalisés[5] - [7]. De plus, la théorie semblait indiquer que les conditions qui permettaient le déplacement de caractères étaient limitées. Cet examen a contribué à motiver les avancées théoriques et méthodologiques ainsi que le développement d'un cadre plus rigoureux pour tester le déplacement des caractères[7].
Six critères ont été développés pour établir le déplacement de caractères comme mécanisme de différence entre espèces sympatriques[8] - [9]. Ceux-ci sont :
- les différences entre les taxons sympatriques sont plus importantes que celles qu'on pourrait obtenir par le hasard ;
- les différences dans les états des caractères sont liées aux différences dans l'utilisation des ressources ;
- les ressources étant limitées dans la nature, la compétition interspécifique pour ces ressources est fonction de la similitude des caractères ;
- la distribution des ressources est la même en sympatrie et en allopatrie, de sorte que les différences dans les états des caractères ne sont pas dues à des différences dans la disponibilité des ressources ;
- les différences doivent avoir évolué in situ ;
- les différences doivent être basées sur la génétique[8].
La mise à l'essai rigoureuse de ces critères nécessite une approche synthétique, combinant des domaines de recherche comme l'écologie des communautés, la morphologie fonctionnelle, l'adaptation, la génétique quantitative et la systématique phylogénétique[5]. Bien qu'il ne soit pas souvent possible de satisfaire aux six critères dans une seule étude portant sur le déplacement de caractères, ils fournissent le contexte nécessaire à la recherche sur le déplacement de caractères[5] - [7].
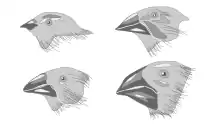
Le déplacement des caractères s'est révélé être un facteur majeur responsable de la taille du bec chez les pinsons des îles Galápagos et des îles Hawaï[10].
Exemples
Des études ont été menées dans une grande variété de taxons — quelques groupes ayant contribué de manière disproportionnée à la compréhension du déplacement des caractères : mammifères carnivores, pinsons des Galápagos, lézards anoles insulaires, épinoches à trois épines et escargots[5].
Oiseaux
Dans l'explication initiale du déplacement de caractères, de nombreux exemples présentés comme preuves potentielles du déplacement du caractères étaient des observations entre plusieurs paires d'oiseaux. Ces exemples incluent les sittelles rocheuses d'Asie, les méliphagidés "mange-miel" du genre Myzantha d'Australie, les perroquets australiens, les puffins des îles du Cap-Vert, les gobe-mouches de l'archipel Bismarck, et bien sûr les pinsons de Darwin dans les Galápagos[1]. David Lack a découvert que lorsque les deux espèces Geospiza fortis et Geospiza fuliginosa étaient présentes ensemble sur de grandes îles, elles pouvaient être distinguées sans équivoque par la taille de leur bec[11]. Cependant, lorsque l'une ou l'autre se trouve seule sur une île plus petite, la taille du bec était de taille intermédiaire par rapport au moment où les deux coexistaient[11]. De même, Peter et Rosemary Grant ont découvert qu'une population insulaire de Geospiza fortis a divergé en taille de bec (en raison d'une mortalité élevée) par rapport à son concurrent G. magnirostris au bout d'une année avec peu de nourriture, apparemment en raison de l'augmentation de la compétition pour des plus grosses graines dont se nourrissaient les deux espèces[12]. La plupart des études sur le déplacement des caractères se concentrent sur les différences morphologiques des caractères impliqués dans l'alimentation plutôt que sur celles liées à l'utilisation de l'habitat. Cependant, des comparaisons de l'utilisation du micro-habitat et des adaptations morphologiques des sittelles rocheuses de l'Ouest et de l'Est indiquent que ces deux espèces présentent une ségrégation de niche spatiale en plus d'une ségrégation de niche trophique (niche alimentaire)[13].
Il est souvent supposé que les espèces étroitement apparentées sont plus susceptibles d'être en compétition que les espèces apparentées plus éloignées, et par conséquent, de nombreux chercheurs étudient le déplacement des caractères entre les espèces du même genre[5]. Bien que le déplacement de caractères ait été initialement débattu dans le contexte d'espèces très proches, des preuves suggèrent que même les interactions entre des espèces éloignées peuvent entraîner un déplacement de caractères. Les pinsons et les abeilles des Galápagos en sont de bons exemples[14]. Deux espèces de pinsons (G. fuliginosa et G. difficilis) exploitent plus de nectar de fleurs sur les îles où l'abeille charpentière blonde (Xylocopa darwini) est absente que sur les îles où elle est présente. Les individus de pinsons qui récoltent le nectar sont plus petits que les membres de la même espèce qui ne le font pas[14].
Reptiles
Le lézard du genre Anolis sur les îles des Caraïbes a également fait l'objet de nombreuses études sur le rôle de la compétition et du déplacement de caractères dans la structure des communautés[15]. Les îles des Petites Antilles ne peuvent accueillir que des espèces d'Anolis de différentes tailles, et l'importance relative du déplacement des caractères par rapport à la taille au moment de la colonisation pour déterminer le succès de l'invasion a été étudiée et débattue.
Amphibiens
Les salamandres des Appalaches Plethodon hoffmani et P. cinereus ne présentent aucune différence morphologique, habitudes alimentaires ou différences d'exploitation de l'utilisation des ressources parmi les populations allopatriques ; lorsque l'espèce est présente en sympatrie cependant, elles présentent une différenciation morphologique associée à une ségrégation dans la taille des proies[16]. Lorsque ces deux espèces coexistent, P. hoffmani a une mâchoire à fermeture plus rapide requise pour les proies plus grosses, et P. cinereus a une mâchoire à fermeture plus lente et plus forte pour les proies plus petites. D'autres études ont trouvé des espèces de salamandres du genre Plethodon qui démontrent un déplacement de caractères par interférence comportementale agressive plutôt que par exploitation[17]. En d'autres termes, le déplacement de caractères morphologiques entre les deux espèces est dû à une interaction agressive entre elles plutôt qu'à l'exploitation de différentes ressources alimentaires.
Mollusques
Sur l'île d'Okinawa, l'espèce d'escargot Satsuma largillierti vit dans la moitié est de l'île, tandis que Satsuma eucosmia vit dans la moitié ouest. Les deux populations se chevauchent en sympatrie le long du milieu de l'île, où la longueur du pénis de l'espèce diffère considérablement là où elles se rencontrent en sympatrie[18]. Les longueurs de pénis des escargots présentent une divergence, suggérant un déplacement du caractère reproducteur de ce trait[19].
Poissons
Les épinoches à trois épines (Gasterosteus spp.) dans les lacs postglaciaires de l'ouest du Canada ont grandement contribué aux récentes recherches sur le déplacement des caractères[20] - [21]. Les observations des populations naturelles et les expériences en laboratoire montrent que lorsque deux espèces récemment apparues dans l'évolution se rencontrent dans un même lac, deux morphologies sont positivement sélectionnées : une forme limnétique qui se nourrit en eau libre et une forme benthique qui se nourrit au fond du lac. Elles diffèrent par leur taille, leur forme, le nombre et la longueur des branchies, et ces différences sont liées à des divergences dans leur alimentation. Les hybrides entre les deux formes sont contre-sélectionnés. Lorsqu'une seule espèce habite un lac, ce poisson présente une morphologie intermédiaire. Des études sur d'autres espèces de poissons ont montré des schémas de sélection similaires pour les morphologies benthiques et limnétiques[5], ce qui peut également conduire à une spéciation sympatrique[22].
Mammifères
Les espèces introduites ont également fourni des "expériences naturelles" récentes pour étudier la rapidité avec laquelle le déplacement des caractères peut affecter le changement évolutif[5]. Lorsque le vison américain (Mustela vison) a été introduit dans le nord-est de la Biélorussie, le vison indigène européen (Mustela lutreola) a augmenté de taille et le vison introduit a diminué de taille[23]. Ce déplacement de caractères a été observé dans une étude ayant porté sur dix ans, démontrant que la compétition peut entraîner des changements évolutifs rapides.
Voir aussi
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Character displacement » (voir la liste des auteurs).
Références
- (en) W. L. Brown et E. O. Wilson, « Character Displacement », Systematic Biology, vol. 5, no 2, , p. 49–64 (ISSN 1063-5157, DOI 10.2307/2411924, lire en ligne, consulté le )
- Thierry Lodé "La guerre des sexes chez les animaux" 2006 Eds Odile jacob, Paris (ISBN 2-7381-1901-8)
- (en) Meyer A, « Phylogenetic Relationships and Evolutionary Processes in East African Cichlid Fishes », sur Trends in ecology & evolution, (PMID 21236169, consulté le )
- (en) Noor Ma, « Reinforcement and Other Consequences of Sympatry », sur Heredity, (PMID 10620021, consulté le )
- (en) Tamar Dayan et Daniel Simberloff, « Ecological and community-wide character displacement: the next generation », Ecology Letters, vol. 8, no 8, , p. 875–894 (ISSN 1461-0248, DOI 10.1111/j.1461-0248.2005.00791.x, lire en ligne, consulté le )
- (en) P. R. Grant, « Convergent and divergent character displacement », Biological Journal of the Linnean Society, vol. 4, no 1, , p. 39–68 (ISSN 0024-4066, DOI 10.1111/j.1095-8312.1972.tb00690.x, lire en ligne, consulté le ).
- (en) Losos Jb, « Ecological Character Displacement and the Study of Adaptation », sur Proceedings of the National Academy of Sciences of the United States of America, (PMID 10823930, consulté le )
- (en) Taper Ml et Case Tj, « MODELS OF CHARACTER DISPLACEMENT AND THE THEORETICAL ROBUSTNESS OF TAXON CYCLES », sur Evolution; international journal of organic evolution, (PMID 28564035, consulté le )
- (en) Schluter D et McPhail Jd, « Ecological Character Displacement and Speciation in Sticklebacks », sur The American naturalist, (PMID 19426066, consulté le )
- Dolph Schluter, « Character Displacement and the Adaptive Divergence of Finches on Islands and Continents », The American Naturalist, vol. 131, no 6, , p. 799–824 (ISSN 0003-0147, DOI 10.1086/284823, lire en ligne, consulté le )
- (en) David Lack, Darwin's Finches, Oxford University Press,
- (en) Grant Pr et Grant Br, « Evolution of Character Displacement in Darwin's Finches », sur Science (New York, N.Y.), (PMID 16840700, consulté le )
- Yousefi, M., Kaboli, M., Eagderi, S. et Mohammadi, A., « Micro-spatial separation and associated morphological adaptations in the original case of avian character displacement. », Ibis, vol. 159, no 4, , p. 883–891 (DOI 10.1111/ibi.12505)
- Dolph Schluter, « Character Displacement between Distantly Related Taxa? Finches and Bees in the Galapagos », The American Naturalist, vol. 127, no 1, , p. 95–102 (ISSN 0003-0147, DOI 10.1086/284470, lire en ligne, consulté le )
- (en) Losos Jb, « A PHYLOGENETIC ANALYSIS OF CHARACTER DISPLACEMENT IN CARIBBEAN ANOLIS LIZARDS », sur Evolution; international journal of organic evolution, (PMID 28567973, consulté le )
- (en) Adams Dc et Rohlf Fj, « Ecological Character Displacement in Plethodon: Biomechanical Differences Found From a Geometric Morphometric Study », sur Proceedings of the National Academy of Sciences of the United States of America, (PMID 10760280, consulté le )
- (en) Dean C. Adams, « Character Displacement Via Aggressive Interference in Appalachian Salamanders », Ecology, vol. 85, no 10, , p. 2664–2670 (ISSN 1939-9170, DOI 10.1890/04-0648, lire en ligne, consulté le )
- (en) Kameda Y et Kawakita A, « Reproductive Character Displacement in Genital Morphology in Satsuma Land Snails », sur The American naturalist, (PMID 19298185, consulté le )
- (en) Carl T. Bergstrom et Lee Alan Dugatkin, Evolution, W. W. Norton & Company, , 2e éd., p. 508-509
- (en) Dolph Schluter, « Adaptive Radiation in Sticklebacks: Size, Shape, and Habitat Use Efficiency », Ecology, vol. 74, no 3, , p. 699–709 (ISSN 1939-9170, DOI 10.2307/1940797, lire en ligne, consulté le )
- (en) Dolph Schluter, « Adaptive Radiation in Sticklebacks: Trade-Offs in Feeding Performance and Growth », Ecology, vol. 76, no 1, , p. 82–90 (ISSN 1939-9170, DOI 10.2307/1940633, lire en ligne, consulté le )
- (en) Barluenga M et Stölting Kn, « Sympatric Speciation in Nicaraguan Crater Lake Cichlid Fish », sur Nature, (PMID 16467837, consulté le )
- (en) V. Sidorovich, H. Kruuk et D. W. Macdonald, « Body size, and interactions between European and American mink (Mustela lutreola and M. vison) in Eastern Europe », Journal of Zoology, vol. 248, no 4, , p. 521–527 (ISSN 1469-7998, DOI 10.1111/j.1469-7998.1999.tb01051.x, lire en ligne, consulté le )