Anaschisma
Anaschisma est un genre éteint et fossile de grands amphibiens temnospondyles. L'espèce type est Anaschisma browni et en 2022, le genre est resté montypique.

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Super-classe | Tetrapoda |
| Classe | Amphibia |
| Ordre | † Temnospondyli |
| Sous-ordre | † Stereospondyli |
| Famille | † Metoposauridae |
- Borborophagus Branson & Mehl, 1929
- Buettneria Affaire, 1922
- Kalamoiketer Branson & Mehl, 1929
- Koskinonodon Branson & Mehl, 1929
- Anaschisma brachygnatha Branson, 1905
- Borborophagus wyomingensis Branson & Mehl, 1929
- Buettneria calgariensis Vert, 1954
- Buettneria howardensis Sawin, 1944
- Buettneria perfecta Affaire, 1922
- Kalamoiketer pinkleyi Branson & Mehl, 1929
- Koskinonodon perfectus Affaire, 1922
- Koskinonodon princeps Branson & Mehl, 1929
Présentation
Ces animaux font partie de la famille des Metoposauridae, qui remplissait les niches prédatrices ressemblant à des crocodiles à la fin du Trias[1]. Il avait un grand crâne d'environ 62 cm de long[2], et ayant peut-être atteint 3 m de long[3]. C'était un chasseur embusqué, prenant tout ce qui était assez petit pour tenir dans ses énormes mâchoires[3]. Il était très courant au Trias supérieur (âge norien) dans ce qui est aujourd'hui le sud-ouest américain[4].
Histoire de la découverte
Anaschisma a été nommé en 1905 par le géologue et paléontologue Branson (d) (1877-1950) à partir de deux crânes de Metoposauridae de la formation Popo Agie dans le Wyoming. L'espèce type, Anaschisma browni, a été inventée pour le crâne UC 447, tandis qu'une deuxième espèce nominale, Anaschisma brachygnatha, a été érigée pour le crâne UC 448[5]. Moodie (1908) considérait A. brachygnatha comme un synonyme junior d'A. browni, bien que Branson et Mehl (1929) aient retenu les deux espèces comme distinctes[6] - [7]. Colbert et Imbrie (1956) ont établi une relation de synonymie entre l'anaschisma et le genre Eupelor du Supergroupe de Newark, mais l'ont conservé comme une espèce Eupelor valide endémique attestée dans la Formation de Popo Aggie[8]. Chowdhury (1965) considérait l'anaschisma comme synonyme du Metoposaurus et a fusionné tous les métoposauridés nord-américains du Chinle et du Dockum dans l'A. browni[9].
Hunt (1989) a restauré l'entité d'Anaschisma sous une forme avancée ou hautement dérivée[4]. Certains spécimens attribués à Anaschisma de la formation de Redonda ont été renommés Apachesaurus par Hunt (1993)[10] - [11].
Problèmes de terminologie autour du genre Koskinonodon

Le genre Koskinonodon était autrefois nommé Buettneria par Case en 1922, mais en 2007, B.D. Mueller s'est rendu compte que le nom Buettneria avait déjà été donné à un grillon de brousse de la République du Congo par Karsch (1889), il a donc fait du genre Koskinonodon le premier nom disponible sans préoccupation pour le temnospondyle[12]. Lucas et al. (2007), cependant, ont demandé au CINZ de supprimer le nom de Karsch en faveur du nom de Case, citant des preuves que le nom d'amphibien est beaucoup plus connu et largement utilisé (les auteurs ont cité 75 utilisations du nom dans la littérature et les livres scientifiques au cours des dernières 85 ans), et que le nom de criquet de brousse avait rarement été utilisé dans la littérature technique[13]. Cependant, l'avis 2255 du CINZ publié en 2010 a rejeté la requête[14].
Des fossiles connus de Koskinonodon ont été trouvés aux États-Unis, en particulier dans la formation de Chinle au Parc National de la forêt pétrifiée et à la carrière de Placerias en Arizona, la formation de Garita Creek au centre du Nouveau-Mexique (la carrière de Lamy), à une partie de la forêt pétrifiée du nord du Nouveau-Mexique, dans la formation Bluewater Creek de l'ouest du Nouveau-Mexique, dans la formation New Oxford de Pennsylvanie, dans la formation Tecovas de l'ouest du Texas et dans la formation Popo Agie du Groupe Chugwater du Wyoming[7] - [15].
La synonymie de Koskinonodon avec Anaschisma a commencé lorsque Romer (1947) a proposé de considérer Anaschisma comme un synonyme senior des genres Buettneria, Koskinonodon et Borborophagus[15]. Cet avis a été suivi par Gee et al. (2019), où les auteurs ont redécrit les holotypes des deux espèces nominales du genre Anaschisma, puis ont posé la synonymie des genres Koskinonodon, Buettneria et Borborophagus avec Anaschisma[16].
Description
Crâne
Le crâne d'Anaschisma diffère des crânes d'autres organismes étroitement apparentés sur quelques points clés. Il est globalement plus large et présente des orbites très antérieures et orientées vers l'avant, plus que ses contemporains[6]. De plus, le crâne a des canaux visqueux proéminents, qui sont utilisés pour transporter le mucus, ainsi que de grandes narines externes[17]. De plus, la mâchoire supérieure est relativement faible et mince, utilisée uniquement pour tenir les dents[17]. Leurs grandes mâchoires auraient pu contenir plusieurs dents à la fois, peut-être même plus de cent de chaque côté des mâchoires supérieure et inférieure, mais le nombre réel varie constamment au cours de la vie de l'animal en raison de causes naturelles telles que les combats, l'alimentation, la maladie, etc.[17] La taille du crâne est l'un des traits les plus déterminants du genre Anaschisma[17].
D'autres traits crâniens plus mineurs caractérisent également Anaschisma comme l'allongement du lacrymal, le raccourcissement du préfrontal, la réduction de l'interclavicule, et, le plus caractéristique, le lacrymal central entrant dans la marge de l'orbite[3]. Alors que le préfrontal raccourci est une caractéristique de la famille des Metoposauridae, il est encore plus court chez Anaschisma[1]. L'interclavicule est réduite dans la mesure où elle comporte de nombreuses fosses hexagonales ainsi que des rainures et des crêtes[1]. Le crâne d’Anaschisma est également recouvert de cette ornementation réticulée[18]. Certains chercheurs pensent qu’Anaschisma a un processus postérieur plus court de l'interclavicule, ce qui peut le différencier d'autres espèces étroitement apparentées, tandis que d'autres pensent qu'il n'y a pas suffisamment d'informations pour faire cette distinction. Anaschisma a également des opisthotiques ossifiés, les plus postérieurs des os entourant l'oreille interne[19].
Dentition
Anaschisma avait des dents acérées et pointues pour attraper et tuer des proies. Ils avaient des dents marginales ainsi que des dents plus grandes sur le palais, en particulier des dents palatines et ectoptérygoïdiennes[1]. Ils avaient deux types de base : les grands avec des rainures peu profondes et les petits avec des rainures plus profondes. Ces rainures s'étendent dans le sens de la longueur des dents et aident les animaux à attraper des proies[18]. Les dents sont allongées labio-lingualement à leur base et opposées, médio-distalement, à leur pointe. Au milieu, ils ne sont pas allongés dans les deux sens mais plutôt circulaires[18]. Ces adaptations dentaires ont amélioré la capacité d’Anaschisma à capturer des proies ; les dents sont optimisées pour percer les proies et ne pas leur permettre de s'échapper, résister à la force de flexion appliquée par l'organisme en difficulté et propager des fissures dans les parties dures de l'objet, comme les os, permettant une alimentation et une digestion plus faciles[18].
Membres inférieurs
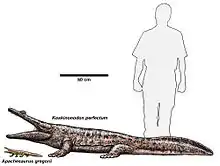
Anaschisma avait une queue large, en forme de coin et puissante pour l'aider à nager, à chasser et probablement à se défendre. Il n'était pas long comme les crocodiles d'aujourd'hui, mais plus probablement court et fort pour lui permettre de sortir rapidement de sa cachette et de capturer des proies avant qu'elles ne s'échappent[3]. Les jambes d’Anaschisma affichent une posture sinueuse et des jambes courtes avec quatre doigts sur le devant et cinq sur les membres arrière. Anaschisma a probablement passé beaucoup de temps immobile, à attendre une proie, à laquelle ces pattes courtes étaient probablement issues d'un processus d'adaptation[5].
Aspects paléobiologiques

Anaschisma, pour chasser, s'allonge au fond d'un marais peu profond, attendant qu'un poisson, un crustacé, un amphibien plus petit ou même un jeune phytosaure se promène[3]. Lorsqu'il a repéré sa proie il utilisait ses énormes mâchoires pour les engloutir[3]. Quelques adaptations particulières suggèrent que Koskinonodon avait ce mode de vie aquatique. Premièrement, ils avaient des lignes latérales formées par les sillons sensoriels. Ceux-ci sont utiles pour détecter les changements de pression de l'eau causés par les mouvements de nage des organismes à proximité. Leurs membres tentaculaires étaient également adaptés à l'eau. Ils ne se déplaceraient pas rapidement ou efficacement sur terre, bien qu'ils l'aient peut-être fait pour trouver un autre bassin d'eau avec plus de nourriture ou d'autres ressources. Des fosses communes ont été découvertes, que l'on pense être le résultat d'un groupe de ces animaux se rassemblant dans une piscine d'eau flétrie pendant une sécheresse et tous périssant parce que l'eau n'a jamais été réapprovisionnée[17].
Aspects paléoécologiques
Ces amphibiens vivaient principalement à la fin du Trias ; au moment où le Jurassique a commencé, la plupart des temnospondyles, y compris Anaschisma, avaient disparu[20]. Il est probable qu'ils se soient éteints lors de l'événement d'extinction du Trias-Jurassique, avec la majorité des autres grands amphibiens, la classe des Conodontes et 34% de tous les genres marins. On n'a pas encore définitivement départagé toutes les causes de cette extinction massive, dans la mesure où les diverses théories incluent d'énormes éruptions volcaniques, le changement climatique, l'acidification océanique et même un autre impact d'astéroïde. On sait cependant que plus de la moitié des espèces vivant sur Terre à cette époque se sont éteintes à cause de cet événement[21]. Anachisma, identifié en 1931 par Case, vivait aux côtés de nombreux autres amphibiens plus petits, et ses fossiles se trouvent également couramment avec des fossiles de phytosaures[17] - [22]. Les meilleures conditions de fossilisation se produisent dans les vallées fluviales ou les plaines inondables où se produisent des dépôts, et cet animal vivait probablement dans des habitats marécageux peu profonds similaires. Anaschisma demeure célèbre pour être attesté par des fossiles extrêmement bien conservés, et souvent découverts en groupes[17].
Voir aussi
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
Références
- (en) Stephen L. Brusatte, Richard J. Butler, Octávio Mateus et J. Sébastien Steyer, « A new species of Metoposaurus from the Late Triassic of Portugal and comments on the systematics and biogeography of metoposaurid temnospondyls », Journal of Vertebrate Paleontology, vol. 35, no 3, , e912988 (ISSN 0272-4634, DOI 10.1080/02724634.2014.912988, lire en ligne, consulté le )
- (en) Adrian P. Hunt et Spencer G. Lucas, « Taxonomy and Stratigraphic Distribution of Late Triassic Metoposaurid Amphibians from Petrified Forest National Park, Arizona », Journal of the Arizona-Nevada Academy of Science, vol. 27, no 1, , p. 89–96 (ISSN 0193-8509, lire en ligne, consulté le ).
- (en) « Palaeos Vertebrates Temnospondyli: Trematosauria: Metaposaurs », sur palaeos.com (consulté le )
- (en) Hunt A. P., « Comments on the taxonomy of North American metoposaurids and a preliminary phylogenetic analysis of the family Metoposauridae », Lucas, SG; Hunt, AP (eds.). Dawn of the age of dinosaurs in the American Southwest., Albuquerque, New Mexico Museum of Natural History, , p. 292-300 (lire en ligne)
- E. B. Branson, « Structure and Relationships of American Labyrinthodontidæ », The Journal of Geology, vol. 13, no 7, , p. 568–610 (ISSN 0022-1376, DOI 10.1086/621258, lire en ligne, consulté le )
- (en) Roy L. Moodie, « The lateral line system in extinct amphibia », Journal of Morphology, vol. 19, no 2, , p. 511–540 (ISSN 0362-2525 et 1097-4687, DOI 10.1002/jmor.1050190206, lire en ligne, consulté le )
- Branson, E. B. & Mehl, M. G. 1929. Triassic amphibians from the Rocky Mountain region. The University of Missouri Studies, 4, 154–253.
- (en) Colbert, Edwin Harris et Imbrie, John, « Triassic metoposaurid amphibians », Bulletin of the American Museum of National History, vol. 110, , p. 405-452 (lire en ligne)
- T. Roy Chowdhury et Prasanta Chandra Mahalanobis, « A new metoposaurid amphibian from the upper Triassic Maleri formation of Central India », Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, vol. 250, no 761, , p. 1–52 (DOI 10.1098/rstb.1965.0019, lire en ligne, consulté le )
- (en) Tomasz Sulej, « Species discrimination of the Late Triassic temnospondyl amphibian Metoposaurus diagnosticu », Acta Palaeontologica Polonica, vol. 47, no 3, (lire en ligne)
- Hunt, A.P. 1993. Revision of the Metoposauridae (Amphibia: Temnospondyli) and description of a new genus from Western North America. In: M. Morales (eds.), Aspects of Mesozoic Geology and Paleontology of the Colorado Plateau. Museum of Northern Arizona Bulletin 59: 67–97.
- Bill D. Mueller, « Koskinonodon Branson and Mehl, 1929, a replacement name for the preoccupied temnospondyl Buettneria Case, 1922 », Journal of Vertebrate Paleontology, vol. 27, no 1, , p. 225–225 (ISSN 0272-4634, DOI 10.1671/0272-4634(2007)27[225:KBAMAR]2.0.CO;2, lire en ligne, consulté le )
- Lucas, S. G., Rinehart, L. F. et Spielmann, J. A., « Comments on the proposed conservation of Buettneria Case, 1922 (Amphibia) 2 (Case 3420) », Bulletin of Zoological Nomenclature., vol. 65, no 3, , p. 218-219 (lire en ligne)
- « OPINION 2255 (Case 3420) Buettneria Case, 1922 (Amphibia): generic name not conserved | International Commission on Zoological Nomenclature », sur web.archive.org, (consulté le )
- Romer, A. S. 1947. Review of the Labyrinthodontia. Bulletin of the Museum of Comparative Zoology, 99, 1–368.
- Bryan M. Gee, William G. Parker et Adam D. Marsh, « Redescription of Anaschisma (Temnospondyli: Metoposauridae) from the Late Triassic of Wyoming and the phylogeny of the Metoposauridae », Journal of Systematic Palaeontology, vol. 18, no 3, , p. 233–258 (ISSN 1477-2019, DOI 10.1080/14772019.2019.1602855, lire en ligne, consulté le )
- (en) « Anaschisma articles - Encyclopedia of Life », sur eol.org (consulté le )
- (en) Lawrence H. Tanner, Justin A. Spielmann et Spencer G. Lucas, The Triassic System: New Developments in Stratigraphy and Paleontology: Bulletin 61, New Mexico Museum of Natural History and Science, (lire en ligne)
- Rinehart, Larry F; Lucas, Spencer G (2013). "The functional morphology of dermal bone ornamentation in temnospondyl amphibians". In Tanner, Lawrence H.; Justin A. Spielmann; Spencer G. Lucas (eds.). The Triassic System: New Developments in Stratigraphy and Paleontology. Bulletin of the New Mexico Museum of Natural History and Science. Vol. 61. pp. 524–532.
- McHugh, Julia Beth (2012). Temnospondyl ontogeny and phylogeny, a window into terrestrial ecosystems during the Permian-Triassic mass extinction (PhD thesis). Graduate College of the University of Iowa.
- (en) « end-Triassic extinction | Evidence & Facts | Britannica », sur www.britannica.com (consulté le )
- (en) William G. Parker et Jeffrey W. Martz, « The Late Triassic (Norian) Adamanian–Revueltian tetrapod faunal transition in the Chinle Formation of Petrified Forest National Park, Arizona », Earth and Environmental Science Transactions of The Royal Society of Edinburgh, vol. 101, nos 3-4, , p. 231–260 (ISSN 1755-6929 et 1755-6910, DOI 10.1017/S1755691011020020, lire en ligne, consulté le )