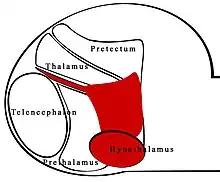
Zona limitans intrathalamica
La zona limitans intrathalamica (ZLI) est un compartiment de restriction de lignée et une limite de développement primaire dans le cerveau antérieur des vertébrés (analogue au cerveau humain) servant de centre de signalisation et de frontière restrictive entre deux régions du diencéphale.
La signalisation Sonic hedgehog (shh) de la ZLI est cruciale dans le développement du diencéphale[1], qui se développe dans le thalamus, le pretectum et les structures tegmentales antérieures[2]. la ZLI avec le préthalamus et le thalamus constituent le territoire moyen-diencéphalique (MDT)[3].
Découverte
Les frontières de restriction de la lignée cellulaire, à travers lesquelles les cellules en réplication ne peuvent pas migrer, ont d'abord été découvertes chez les invertébrés, où l'expression de divers gènes Hox dans chaque compartiment confère une différenciation des segments observables dans le corps adulte de Drosophila melanogaster. Des structures analogues ont été découvertes dans le cerveau en développement des vertébrés. Les rhombomères, qui descendent de l' embryon à partir du cerveau postérieur, contiennent des limites claires et expriment chacun divers gènes Hox nécessaires à la différenciation des structures dans le corps. Des régions plus antérieures du cerveau ont été examinées à la recherche d'autres limites de restriction de lignée cellulaire, et de multiples limites potentielles continuent d'être étudiées (voir Limites de développement)[4].
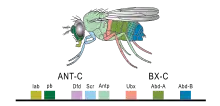
L'importance de ces compartiments en tant que centres de signalisation locaux (des zones qui influencent chimiquement les tissus environnants) a été élucidée en observant d'abord l' expression différentielle des gènes Hox dans divers compartiments et ensuite en observant le mutant D. melanogaster et les changements phénotypiques (physiques) correspondants[4].
la ZLI a été découvert pour la première fois chez le poussin à l'aide d'expériences d'explantation et de marquage de lignées. Dans les expériences d'explantation, les cellules de la région qui deviendra la ZLI, le préthalamus et le thalamus du poussin ont été prélevées et placées dans des cultures séparées ; les cellules ont continué à croître et à conserver leur identité (la ZLI a commencé à transcrire shh, contrairement au préthalamus et au thalamus ). La nécessité de la ZLI et de son expression shh correspondante a été mise en évidence par le manque de marqueurs génétiques thalamiques et préthalamiques en culture (voir Signalisation)[5]. Ces expériences ont confirmé la ZLI en tant que centre de signalisation. Dans les expériences de marquage de lignées, les cellules étaient génétiquement marquées, de sorte que chaque fois qu'une cellule marquée se répliquait, sa descendance était également marquée. Les cellules qui ont été marquées dans la ZLI en développement et leur descendance sont restées limitées à la zone. Ces expériences ont démontré la ZLI comme une limite de restriction de lignée cellulaire[6].
En plus d'être un frontère, la ZLI est également un compartiment avec des limites de restriction de lignée cellulaire séparées à la fois antérieures et postérieures d'une section d'expression chut. L'importance de la ZLI a été une fois de plus confirmée par l' expression ectopique de shh dans d'autres régions du cerveau antérieur, connues sous le nom de prosencéphale au cours du développement (à la fois le télencéphale et le diencéphale ), induisant une région de type ZLI qui induit le destin thalamique[7].
Limites de développement
Au cours du développement chez les vertébrés et les invertébrés, les limites de restriction de la lignée cellulaire et les centres de signalisation sont formés afin d'assurer une différenciation appropriée du corps. Les signaux chimiques, comme Shh dans la ZLI, sont souvent libérés de ces limites et compartiments dans un gradient de concentration (les produits chimiques sont à des concentrations beaucoup plus élevées plus près de la source) et confèrent une identité aux régions flanquantes. D'autres gènes exprimés de manière différentielle dans ces régions flanquantes aident à assurer une différenciation appropriée (voir Signalisation).
De nombreuses limites de développement ont été étudiées : dans le cerveau antérieur uniquement, les limites confirmées de restriction de la lignée cellulaire sont la limite palléale-sous-pallée (PSB) divisant le télencéphale dorsal et ventral, la limite diencéphale-mésencéphale (DMB) postérieure à la ZLI et la ZLI . la ZLI, comme chaque rhombomère, sert de compartiment indépendant qui confère l'identité du diencéphale dans les régions antérieure et postérieure. D'autres limites de développement servent de limites de restriction de lignée cellulaire mais pas de centres de signalisation, tandis que d'autres sont des centres de signalisation vers et à partir desquels les cellules peuvent migrer. Malgré les découvertes de limites et de compartiments de restriction de lignée cellulaire dans le cerveau, de nombreuses régions étudiées ont été réfutées en tant que limites segmentaires. Ces zones ont un potentiel en tant que centres de signalisation, qui ont une influence sur le développement des tissus voisins[8].
Ces limites ont une grande influence sur les autres régions du cerveau : le placement de la ZLI affecte non seulement la taille des régions adjacentes mais aussi la taille du télencéphale . Un décalage postérieur de la ZLI permet d'allouer plus de cellules au télencéphale . Il en va de même pour les autres limites du développement dans le cerveau et dans tout le corps : les changements de limites responsables de l'affectation d'une certaine quantité de tissu à une certaine fonction entraînent des changements drastiques dans la structure adulte. Ces limites sont d'une importance cruciale pour une différenciation appropriée.
Formation
Motif d'axe initial
Après la gastrulation, l' embryon est complètement indifférencié et nécessite de nombreux signaux différents pour initier une différenciation appropriée du corps. Le haut (plaque de toit, sur la face dorsale de l'embryon) et le bas (plaque de sol, sur la face ventrale) jouent un rôle crucial dans ces premières étapes : chacun agit par une signalisation globale (signalisation dans tout l'embryon) pour la structuration neuronale dorsoventrale . Une fois le développement de l' axe dorsoventral terminé, une signalisation plus locale se produit dans le cerveau en développement : les limites du développement telles que la limite mésencéphale-cerveau postérieur (MHB), les rhombomères et la ZLI aident à l'organisation antéropostérieure. [5].
Émergence de l'expression Shh
Peu de temps après le début de la structuration dorso- ventrale, Shh s'exprime le long de la plaque basale (le bas) de l' embryon, qui fonctionne dans « la ventralisation du tube neural, la promotion de la croissance et de la prolifération, et la formation de l' hypothalamus »[9]. Au fur et à mesure que l' embryon continue de se développer, l' expression Shh caractéristique de la ZLI s'étend dorsalement pour former un coin qui se rétrécit finalement en une bande à environ 22 somites (le nombre de rhombomères développés) ou moins d'un jour chez le poisson zèbre . Bien que l'expression de Shh s'étende dorsalement à partir de la plaque basale, la ZLI est capable de se former même sans la plaque basale ou le tissu mésodermique. Shh coopère avec <i id="mwqQ">dlx2</i> et fezl en avant et IRX3 et dbx1a en arrière (les gènes exprimés varient selon les organismes), qui sont des gènes exprimés dans le préthalamus et le thalamus, respectivement[10].
La ZLI est également caractérisée par une absence de frange lunatique ( lfng ), qui est observable avant même que l' expression Shh ne soit observée dans la ZLI en formation. Cela indique que les cellules sont destinées à la formation de ZLI avant l'induction par Shh (nb Shh]' des signaux de la ZLI et est caractéristique du compartiment pendant et après la formation). Lfng est exprimé très tôt (peu après la gastrulation ) dans la région qui deviendra la ZLI, mais peu de temps après, l'expression diminue par migration des cellules exprimant lfng pour former le coin sans lfng caractéristique de la ZLI en développement. L' expression Shh ne s'étend dorsalement que quelques heures plus tard[5].
Positionnement
Les facteurs influençant la formation et la localisation de la ZLI sont largement étudiés mais toujours contestés. Les différences entre les différents modèles animaux compliquent davantage l'élucidation des voies génétiques, en particulier entre les modèles poussins/mammifères et le poisson zèbre .
Les gènes Wnt (la famille sans ailes) sont cruciaux pour le développement de la ZLI à la fois directement et indirectement dans tous les systèmes animaux. Parallèlement au rôle des gènes Wnt dans la structuration de l' axe antéropostérieur par polarisation de gradient, Wnt8b est exprimé dans la ZLI lui-même et peut aider à guider le mouvement dorsal de l'expression Shh.[11]. Le gradient de polarisation Wnt a été lié à l'induction des gènes de configuration ZLI IRX3 et SIX3, qui bordent la ZLI en arrière et en avant, respectivement. Cependant, ces gènes se sont avérés non essentiels pour la formation de ZLI chez le poisson zèbre et ont été réévalués dans d'autres modèles[12] - [10].
La spécification de la ZLI peut également concerner les tissus ventraux de l' embryon : les plaques préchordale et épichordale, ou le tissu neuroépithélial (voir figure). Les interactions entre ces tissus peuvent être responsables de la récession d'expression de <i id="mw5g">lfng</i> qui permet le mouvement dorsal de l' expression Shh. [13]. Les plaques préchrodales et épichordales spécifiques, caractérisées par l'expression de SIX3 et IRX3, respectivement, peuvent influencer le positionnement de la ZLI plus que les gènes eux-mêmes[5]. La plaque préchrodale borde ventralement le télencéphale, avec la plaque épichrodale postérieure à celui-ci.
Des études sur la formation de la ZLI réalisées chez le poisson zèbre ont révélé l'importance d' OTX2 et d' IRX1 dans le positionnement de la ZLI. L' expression d' OTX2 caractérise le tectum optique en développement, responsable du traitement de la vue. L'expression s'étend vers l' avant et se termine brusquement au ZLI, avec une expression élevée le long de la ligne où Shh est exprimé[14]. Avant la formation de la ZLI, OTX2] est exprimé de manière omniprésente dans tout le cerveau antérieur et commence à reculer vers la position de la ZLI putative. Les expériences où l' expression d'OTX2] était réprimée n'ont montré aucun mouvement dorsal de l' expression de Shh et aucune formation de ZLI[10]. Irx1, qui est analogue à IRX3 chez les oiseaux et les mammifères, est exprimé postérieurement à la ZLI. Des études ont suggéré que bien que OTX2 restreigne positivement la ZLI antérieurement ( Shh ne peut pas être exprimé là où OTX2 ne l'est pas), « IRX1 restreint négativement la ZLI postérieurement[10]. D'autres gènes cruciaux pour la différenciation dans le cerveau, y compris les gènes Fgf responsables de la structuration de la limite mésencéphale-cerveau postérieur (MHB), ont été impliqués dans le positionnement de la ZLI[11].
Les études sur le rôle de la signalisation Shh dans la ZLI ont été difficiles à étudier pendant de nombreuses années, car les mutants manquant d'expression présentent de nombreux déficits de développement, notamment l'absence de diencéphale [15]. Les expériences d'explantation et de marquage de lignées décrites précédemment ont aidé à élucider le rôle de Shh et d'autres gènes dans la différenciation de ces tissus . Plus récemment, le double mutant de souris Shh;Gli3 s'est avéré avoir un diencéphale élargi avec un anneau de Fgf8 et Wnt à la place de la ZLI, indiquant une interaction complexe entre Shh et ces gènes au niveau de la ZLI[16]. Cela indique également que d'autres signaux de structuration sont capables d'établir des domaines de signalisation Fgf8 et Wnt au ZLI en l'absence de Shh et Gli3.
Différenciation après dégradation ZLI
Après différenciation des cellules progénitrices (à un stade précis encore à déterminer), la ZLI et sa restriction de lignée disparaissent, permettant aux cellules de migrer à travers l'ancienne limite et les thalami dorsal et ventral de fusionner en une unité fonctionnelle, comme le montre expériences rétrovirales incapables de se répliquer qui ont marqué les cellules et montré leur migration à travers le diencéphale[11]
Signalisation
Après l'établissement de la ZLI, il a été démontré que shh induit l'expression de marqueurs thalamiques et préthalamiques, respectivement gbx2 et <i id="mwATE">dlx2</i> / nkx2.1 . Cette induction différentielle est très probablement due à l'expression de gènes tels que IRX3 dans le thalamus : des expériences d'expression ectopique ont montré que si IRX3, qui est normalement exprimé dans le thalamus en développement, est exprimé avant la ZLI, alors le préthalamus en développement changera d'identité[13]. Notez que ces gènes qui aident à conférer la compétence shh aident à modeler la ZLI.
La signalisation de la ZLI coopérant avec les marqueurs thalamiques et préthalamiques assure la migration des cellules post-mitotiques (progénitrices neurales) vers la zone du manteau où les cellules s'assemblent en noyaux caractéristiques du thalamus . Ces noyaux sont le mécanisme de relais de l'information du thalamus vers le cortex . Le thalamus lui-même est très diversifié, chaque noyau ayant des morphologies et des physiologies distinctes selon la région du cerveau à laquelle il est connecté. On pense que ces différences proviennent de l'expression différentielle des gènes dans le thalamus et le préthalamus, ce qui permet une structure unique avec de multiples fonctions différentes et séparées une fois que les deux ont fusionné et terminé la croissance et la différenciation[11]. Ainsi, une interaction fonctionnelle entre Shh de la ZLI et le facteur bHLH Her6 (homologue de HES1 ) détermine l'identité neuronale au sein du thalamus : les cellules Her6 positives du préthalamus et du thalamus rostral se différencient en neurones inhibiteurs GABAergiques, tandis que ses cellules négatives deviennent des neurones relais glutamatergiques . Les deux types de cellules dépendent du signal Shh comme déclencheur pour lancer le programme de développement[17].
Références
- (en) Steffen Scholpp et Andrew Lumsden, « Building a bridal chamber: development of the thalamus », Trends in Neurosciences, vol. 33, no 8, , p. 373–380 (PMID 20541814, PMCID PMC2954313, DOI 10.1016/j.tins.2010.05.003, lire en ligne, consulté le ).
- (en) Raquel Garcia-Lopez, Claudia Vieira, Diego Echevarria et Salvador Martinez, « Fate map of the diencephalon and the zona limitans at the 10-somites stage in chick embryos », Developmental Biology, vol. 268, no 2, , p. 514–530 (DOI 10.1016/j.ydbio.2003.12.038, lire en ligne, consulté le ).
- (en) Steffen Scholpp, Isabelle Foucher, Nicole Staudt et Daniela Peukert, « Otx1l, Otx2 and Irx1b establish and position the ZLI in the diencephalon », Development, vol. 134, no 17, , p. 3167–3176 (ISSN 1477-9129 et 0950-1991, PMID 17670791, PMCID PMC7116068, DOI 10.1242/dev.001461, lire en ligne, consulté le ).
- (en) Clemens Kiecker et Andrew Lumsden, « Compartments and their boundaries in vertebrate brain development », Nature Reviews Neuroscience, vol. 6, no 7, , p. 553–564 (ISSN 1471-003X et 1471-0048, DOI 10.1038/nrn1702, lire en ligne, consulté le ).
- (en) Maria Flavia Guinazu, David Chambers, Andrew Lumsden et Clemens Kiecker, « Tissue interactions in the developing chick diencephalon », Neural Development, vol. 2, no 25, , p. 1–15 (PMID 17999760, PMCID 2217525, DOI 10.1186/1749-8104-2-25).
- (en) « A new developmental compartment in the forebrain regulated by Lunatic fringe », Nature Neuroscience, vol. 4, no 7, , p. 683–684 (PMID 11426219, DOI 10.1038/89455).
- (en) « Expression patterns of homeobox and other putative regulatory genes in the embryonic mouse forebrain suggests a neuromeric organization », Trends in Neurosciences, vol. 16, no 11, , p. 472–479 (PMID 7507621, DOI 10.1016/0166-2236(93)90080-6).
- « Compartments and their boundaries in vertebrate brain development », Nature Reviews Neuroscience, vol. 6, no 7, , p. 553–564 (PMID 15959467, DOI 10.1038/nrn1702).
- « Hedgehog signaling from the zona limitans intrathalamica orchestrates patterning of the zebrafish diencephalon », Development, vol. 133, no 5, , p. 855–864 (PMID 16452095, DOI 10.1242/dev.02248).
- (en) S. Scholpp S, I. Foucher, N. Staudt, D. Peukert D et A. Lumsden A, « Otx1l, Otx2, and Irx1b establish and position the ZLI in the diencephalon », Development, vol. 134, no 17, , p. 3167–3176 (PMID 17670791, DOI 10.1242/dev.001461).
- Y. Lim et JA. Golden, « Patterning the developing diencephalon », Brain Research Reviews, vol. 53, no 1, , p. 17–26 (PMID 16876871, DOI 10.1016/j.brainresrev.2006.06.004).
- (en) « Building a bridal chamber: development of the thalamus », Trends Neurosci, vol. 33, no 8, , p. 373–380 (PMID 20541814, PMCID 2954313, DOI 10.1016/j.tins.2010.05.003)
- C. Vieira, A.L.Garda, K. Shimamura et S. Martinez, « Thalamic development induced by Shh in the chick embryo », Developmental Biology, vol. 284, no 2, , p. 351–363 (PMID 16026780, DOI 10.1016/j.ydbio.2005.05.031).
- « A potential role for the OTX2 homeoprotein in creating early 'highways' for axon extension in the rostral brain », Development, vol. 125, no 21, , p. 4273–4282 (PMID 9753681).
- « Differential activities of Sonic hedgehog mediated by Gli transcription factors define distinct neuronal subtypes in the dorsal thalamus », Mechanisms of Development, vol. 120, no 10, , p. 1097–1111 (PMID 14568100, DOI 10.1016/j.mod.2003.09.001).
- (en) Rash et Grove, « Shh and Gli3 regulate formation of the telencephalic-diencephalic junction and suppress an isthmus-like signaling source in the forebrain. », Developmental Biology, vol. 359, no 2, , p. 242–50 (PMID 21925158, PMCID 3213684, DOI 10.1016/j.ydbio.2011.08.026).
- Scholpp S, Delogu A, Gilthorpe J, Peukert D, Schindler S, Lumsden A. Her6 regulates the neurogenetic gradient and neuronal identity in the thalamus. Proc Natl Acad Sci U S A. 2009 Nov 24;106(47):19895-900.