Trichophyton rubrum
Trichophyton rubrum est un champignon filamenteux microscopique[1] rattaché aux Ascomycota. Il n'a pas de forme sexuée connue. Il a une affinité marquée pour la kératine (protéine de la couche cornée de l'épiderme, et qui constitue les ongles, poils et cheveux).
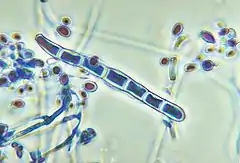
Ce dermatophyte anthropophile (qui infecte l'Homme) est le responsable principal des dermatophytoses des pieds (ou pieds d'athlète, tinea pedis) et des ongles (onychomycose, onyxis, teigne de l'ongle aussi dite tinea unguium). C'est en 2013 le dermatophyte le plus fréquemment isolé dans les laboratoires à partir des mycoses des pieds en Europe et en Amérique du Nord (il y est responsable de plus de 90 % des cas d'onychomycose). Dans certaines régions, comme l'Afrique, il est principalement responsable de la mycose de la peau glabre (herpès circiné, tinea corporis) et des cheveux (teigne, tinea capitis).
Les médecins signalent que ce champignon tend à coloniser un nombre de plus en plus grand d'autres endroits du corps que ce qui a été signalé précédemment[2]. Un ongle en bonne santé y est généralement insensible[2].
Origine
T. rubrum est un parasite obligatoire de l'homme qui est difficilement transmissible aux animaux. Peut-être originaire d'Afrique[3], il s'est ensuite répandu dans le monde entier au XXe siècle.
Étymologie
Le nom scientifique du genre Trichophyton a été construit à partir des racines grecques thríx θριξ−τριχος « cheveu » et phytón φυτον « plante », la référence aux plantes venant du fait que de l'Antiquité gréco-latine jusqu'au milieu du XXe siècle, les champignons étaient classés parmi les plantes ; l'épithète spécifique rubrum vient du latin ruber-rubra-rubrum « rouge » parce qu'en culture sur milieu de Sabouraud, un pigment rouge apparaît.
Castellani isola le dermatophyte en 1909 à Colombo au Sri Lanka (ex-Ceylan) et le nomma Epidermophyton rubrum Castell., Philippine Journal of Science, Section B, Medical Science 5 (2): 203 (1910). Il constituait l'agent principal des mycoses des pieds en Asie. Cette même année 1910, H. Bang, à l'hôpital Saint-Louis, à Paris, l'isola sur deux personnes qui avaient probablement été infectées en Amérique[4] - [5]. Il le nomma Trichophyton purpureum. Ce dermatophyte, apparemment originaire d'Afrique, s'est ensuite répandu dans le monde entier.
En 1910, R. Sabouraud publia le troisième de ses ouvrages sur les maladies du cuir chevelu, consacré aux maladies cryptogamiques et intitulé « Les teignes ». Il y classe les dermatophytes suivant l'aspect de la lésion qu'ils provoquent[6]. Cet ouvrage sera considéré pendant plusieurs décennies comme la référence majeure sur le sujet.
Caractéristiques biologiques
On ne connait pas de forme sexuée (téléomorphe) de Trichophyton rubrum. Il se reproduit sur la gélose de Sabouraud en formant des hyphes mycéliennes et des spores par voie asexuée, on parle de conidies. Ces spores se forment par condensation du cytoplasme à l’intérieur de l'hyphe préexistante puis par fragmentation, on les qualifie d’arthroconidies. Il existe deux types de spores, les macroconidies, qui sont pluricellulaires, et les microconidies, qui sont unicellulaires. Les spores jouent un rôle important dans la propagation du champignon. Elles n'ont pas de besoins nutritionnels exogènes. Ce sont des formes capables de résister aux conditions environnementales adverses et donc de participer à la résistance au traitement antifongique. On considère que les conidies sont la cause des infections primaires de l'homme[7].
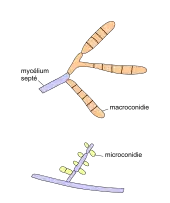
Le diagnostic mycologique peut être réalisé en laboratoire sur une culture de 5 à 10 jours. Le mode de formation des conidies (conidiogenèse) et la structure du mycélium permet de discriminer les différentes espèces de dermatophytes.
- Mycélium à hyphes septés (cloisonnés), filaments en raquettes
- Macroconidies[8] pluricellulaires, à paroi lisse et mince avec plusieurs cloisons (1 à 10), isolées ou en bouquets et de forme variable (en cigare, en saucisse...). La taille est 10 à 85 μm sur 4 à 15 μm.
- Microconidies unicellulaires, piriformes en acladium[n 1] ou rondes. Les microconidies sont l'agent causal primaire des infections de la peau et des ongles.

T. rubrum comme tous les dermatophytes anthropophiles associés à un parasitisme strict (T. violaceum, T. soudanense, M. audouinii, T. schoenleinii) possède :
- une conidiogenèse faible[9] (peu de microconidies, et rares macroconidies),
- une absence d'organe perforateur,
- une dépendance des vitamines et
- une perte de la capacité à dégrader l'urée (test à l'uréase négatif).
En culture, la croissance est rapide. Les conditions optimales de formation des conidies[10], sur une gélose de Sabouraud à dextrose, sont une température de 37 °C, un pH basique de 7,5, sous une atmosphère contenant 10 % de CO2, avec une production maximale au bout de 10 jours.
On peut observer plusieurs types de colonies en culture. Les deux principales sont[11] :
- T. rubrum variété duveteuse (autochtone) le plus souvent isolée en France, formant des colonies plates, blanches et duveteuses, avec un revers coloré d'un pigment rouge brun à violet (lorsque le pH est à 6 - 6,5). Les microconidies sont peu abondantes, disposées en acladium. Les macroconidies sont rarement observées.
- T. rubrum variété poudreuse, correspondant à des souches tropicales d'origine africaine ou asiatique, formant des colonies planes ou plissées, d'aspect poudreux. La conidiogenèse est très abondante : nombreuses microconidies, et macroconidies de calibre régulier (dite en saucisse).
Trichophyton rubrum présente quelques affinités avec Trichophyton interdigitale mais s'en différencie par la pigmentation rouge de ses colonies sur milieu d'épreuve de Sabouraud, ses nombreuses macroconidies et sa faible activité uréasique[12].
Activité kératinolytique
Trichophyton rubrum est un parasite obligatoire de l'homme, spécialisé dans la dégradation de la kératine, une protéine fibreuse, riche en acide aminés à base de soufre (comme la cystéine), susceptibles de former des ponts disulfure[n 2]. Les champignons kératinophiles sont connus pour excréter des protéases (ou protéinases) qui sont capables de dégrader les protéines du milieu en métabolites leur fournissant vraisemblablement du carbone, de l'azote et du soufre, nécessaires à leurs subsistance[13]. Car, pour se nourrir, rappelons que les champignons parasites doivent relâcher dans le milieu des enzymes afin de digérer sur place les macromolécules organiques des tissus de l'hôte et de les transformer en petites molécules qui pourront ensuite être ingurgitées à travers la membrane.
Peu à peu au cours des dernières décennies, les protéinases extracellulaires de Trichophyton rubrum ont été isolées[14]. Fin 2013, la base MEROPS[15] recense 171 protéinases pour T. rubrum. Une étude a montré que plus de la moitié du génome de T. rubrum est composée de protéases, principalement des kératinases[16].
Au niveau des gènes d'endoprotéases, deux familles ont été isolées[17], codant respectivement les subtilases (7 gènes de subtilases SUB de la famille S8 des protéases à sérine, SUB1 à SUB7) et des métalloprotéases (5 gènes de métalloprotéases MEP1 à MEP5 de la famille des fungalysines M36). La production de certaines de ces protéases est déclenchée lorsque le champignon est cultivé sur la kératine comme unique source d'azote. Les subtilases sont des protéases à sérine dont le groupement hydroxyle de la sérine attaque la liaison peptidique. Les métalloprotéases possède un cation métallique qui active une molécule d'eau qui clivera la chaîne peptidique. Une relation directe a pu être établie entre ces protéases et la pathogénicité du dermatophyte.
Monod et al. ont aussi caractérisé des exopeptidases sécrétées : deux aminopeptidases à leucine (LAP) et deux dipeptidyl peptidases (Monod et als[18], 2005) chez T. rubrum.
Ces deux types d'enzymes agiraient en synergie pour produire des oligopeptidases et des acides aminés libres qui pourraient être absorbés par des transporteurs membranaires[19]. Les protéases SUB3 et SUB4 sont actives sur la caséine, le collagène et l'élastine mais surtout la kératine. Toutes les subtilases ne sont cependant pas toutes kératinolytiques ; certaines pourraient avoir des fonctions en rapport avec la réponse immune.
Durant les premiers stades de l'infection, les dermatophytes répondent à la peau en déréprimant certains gènes codant des protéines ou des enzymes, comme les adhésines, lipases, phosphatases, ADNases, protéases non spécifiques et kératinases[20]. Les adhésines, les protéases non spécifiques et les kératinases ont une activité optimale pour un pH acide, qui correspond à l'acidité de la peau[21]. Les peptides, une fois hydrolysés en acides aminés, fournissent une source de carbone, azote et soufre au champignon. La métabolisation de certains acides aminés (comme la glycine), stimule l'alcalinisation du micro-environnement de l'hôte, qui devient alors favorable à l'action de nombreuses kératinases qui sont actives à pH alcalin (Peres et al[22], 2010). Le changement de pH s'accompagne d'une modulation de l'expression des gènes, permettant à T. rubrum de continuer à utiliser les protéines comme source nutritive. Suivant ce modèle, l'adaptation du dermatophyte à un large spectre de pH explique l'installation, le développement et la permanence du champignon dans le tissu de l'hôte.
Répartition et origine
Le séquençage des régions d’ARN ribosomique non codant (ITS[n 3]) a révélé que Trichophyton rubrum possédait des séquences ITS identiques à celles de T. raubitschekii, T. fischeri et T. kanei et très peu différentes de celles de T. soudanense et T. megninii[23]. En s'aidant en outre de cinq marqueurs microsatellites, on peut individualiser environ 30 génotypes au sein du complexe T. rubrum[9]. Ces génotypes peuvent être répartis en trois groupes d'origines géographiques différentes. Le groupe isolé des ongles de pieds ou d'espaces interdigito-plantaires correspond au classique T. rubrum en Europe et à T. kanei en Amérique du Nord. Un deuxième groupe correspond à T. soudanese des teignes du cuir chevelu isolé en Afrique et enfin un groupe d'Asie parasitant la peau et les plis inguinaux (l'aine) correspond à T. raubitschekii.
Si T. rubrum est actuellement le dermatophyte le plus fréquemment isolé dans les laboratoires à partir des mycoses des pieds en Europe et en Amérique du Nord, il n'en a pas toujours été ainsi puisqu'il était quasi absent de ces régions au début du XXe siècle. Les données cliniques enregistrées depuis un siècle mettent en évidence un changement radical du spectre des dermatophytes en Allemagne[24]. Avant la Seconde Guerre mondiale, les dermatophytes dominants étaient Epidermophyton floccosum et Microsporum audouinii (responsable d'épidémie de teignes) alors que Trichophyton rubrum était extrêmement rare. Après la guerre, la part de T. rubrum ne cessa de croître concurremment avec le nombre de patients souffrant d'onychomycose et de pieds d'athlète. On vit ainsi T. rubrum passer de 1 % de l'ensemble des dermatophytes isolés en 1926-1928, à 42 % en 1950 pour atteindre 83 % en 1993. La mise sur le marché d'un traitement à base de griséofulvine a fait quasiment disparaître Epidermophyton floccosum et Microsporum audouinii. Par contre, dans les pays d'Europe méditerranéenne, l'incidence des infections à Microsporum canis a fortement augmenté et ce dermatophyte est devenu dominant pour les cas de tinea capitis chez les enfants.
Son origine africaine ou asiatique est débattue. Suivant une hypothèse de Rippon (1985)[25], l'origine de T. rubrum est à chercher du côté de l'Afrique centrale et de l'Asie orientale. Dans ces foyers originels, il aurait été présent dans les lésions de la peau glabre (mais pas du pied comme aujourd'hui) parmi la population autochtone habituée à marcher nu-pieds. Ce n'est qu'avec l'habitude du port de chaussures fermées dans les villes que la colonisation des espaces interdigitaux plantaires fut favorisée. Les colons et militaires européens auraient ensuite ramené dans leurs chaussures et bottes la nouvelle souche de T. rubrum.
Dans une analyse plus récente, Gräser et al. (2007)[3] suggèrent une origine africaine. Ces auteurs ont collecté plus de 200 isolats de T. rubrum partout dans le monde, dont l'analyse des marqueurs microsatellites a révélé deux populations distinctes. Les génotypes de la première population venaient principalement d'Afrique, alors que ceux de la seconde venaient de partout dans le monde sauf d'Afrique. La plus grande diversité génotypique de la population 1 suggère une origine africaine. Les isolats de la population 1 sont responsables (à plus de 80 %) de mycoses de la peau glabre et des cheveux (tinea corporis et tinea capitis) et très rarement d'infections du pied. Aussi, les auteurs reprennent les considérations de Rippon : les isolats africains viennent des communautés rurales des régions chaudes et humides d'Afrique de l'Ouest où l'habitude de porter des chaussures fermées est peu répandue et où donc les risques de contacter un tinea pedis (pied d'athlète) sont moins élevés. Inversement, les émigrants africains dans les régions tempérées souffrent de tinea pedis et sont infectés par les génotypes non africains du clade 2. Les génotypes locaux semblent donc être mieux adaptés aux conditions environnementales. Les auteurs concluent que l'évolution des populations de T. rubrum est fondamentalement redevable des barrières géographiques.
Clinique
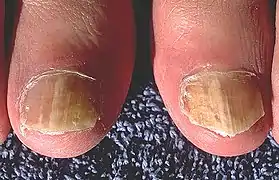
Trichophyton rubrum provoque principalement des lésions interdigito-plantaires (ou « pieds d'athlète ») et des onyxis :
- Intertrigo interdigito plantaire : a pour agents T. rubrum et Epidermophyton floccosum
- Onyxis : les ongles sont contaminés par le bord libre
Ce dermatophyte peut aussi provoquer des lésions onyxis des mains, pachydermie des paumes de main, intertrigo des plis inguinaux (aine) et des épidermophytes circinées éventuellement sur toutes les parties du corps. Il est aussi responsable d'invasions dermiques profondes chez les patients immunodéficients[7].
Notes et références
Notes
- microconidies disposées alternativement d'un côté et de l'autre
- entre deux soufres S de deux cystéines, ce qui consolide la structure
- Internal Transcribed Spacer
Références
- (en) Référence Index Fungorum : Trichophyton rubrum (+ MycoBank)
- Pietro Nenoff, Constanze Krüger, Gabriele Ginter-Hanselmayer, Hans-Jürgen Tietz (2014) Mycology – an update. Part 1: Dermatomycoses: Causative agents, epidemiology and pathogenesis
- Yvonne Gräser, « Microsatellite markers reveal geographic population differentiation in Trichophyton rubrum », Journal of medical microbiology, vol. 56, no Pt 8, , p. 1058-1065 (ISSN 0022-2615, DOI 10.1099/jmm.0.47138-0)
- H. Bang, « Sur une trichophytie cutanée à grands cercles, causée par un dermatophyte nouveau (Trichophyton purpureum Bang.). », Annales de dermatologie et de syphiligraphie. 5e série, vol. 1,
- (en) A.N. Chakraborty, L.S. Ghosh et F. Blank, « Isolation of Trichophyton rubrum (Castellani) Sabouraud 1911, from animals; Cornell Nutrition Conference », Canadian Journal of Comparative Medicine, vol. XVIII, (lire en ligne)
- Raymond Vanbreuseghem, Diagnose et systématique des Dermatophytes. Contribution à la connaissance des Teignes du Congo Belge, Annales de la Société belge de Médecine tropicale, Institut de Médecine Tropicale Prince Léopold, Anvers (dir. Prof. Dubois), (lire en ligne)
- Leng et Tao Liu, Rui Li, Jian Yang, Candong Wei, Wenliang Zhang, Qi Jin, « Proteomic profile of dormant Trichophyton Rubrum conidia », BMC Genomics, vol. 9, no 1, , p. 303 (ISSN 1471-2164, DOI 10.1186/1471-2164-9-303, lire en ligne, consulté le )
- mycoweb
- D. Chabasse, « Les dermatophytes : d’où viennent-ils ? Comment sont-ils devenus des parasites ? », Journal de Mycologie Médicale / Journal of Medical Mycology, vol. 18, no 1, , p. 27-35 (ISSN 1156-5233, DOI 10.1016/j.mycmed.2007.12.002, lire en ligne, consulté le )
- S. Amir Yazdanparast, « Arthroconidia production in Trichophyton rubrum and a new ex vivo model of onychomycosis », Journal of Medical Microbiology, vol. 55, no 11, , p. 1577-1581 (ISSN 0022-2615 et 1473-5644, DOI 10.1099/jmm.0.46474-0, lire en ligne, consulté le )
- Muriel Fromage, S. Houze, M. Miegeville, M. Thellier, « Parasitologie », Annales du Contrôle National de Qualité des Analyses de Biologie Médicale, afssaps,
- (en) Euzéby, Grand dictionnaire illustré de parasitologie médicale et vétérinaire, Lavoisier, (ISBN 9782743018696)
- G. Apodaca, « Purification and characterization of a 27,000-Mr extracellular proteinase from Trichophyton rubrum. », Infection and Immunity, vol. 57, no 10, , p. 3072-3080 (ISSN 0019-9567, lire en ligne, consulté le )
- M. Asahi, « Purification and characterization of major extracellular proteinases from Trichophyton rubrum. », Biochemical Journal, vol. 232, no 1, , p. 139-144 (ISSN 0264-6021, lire en ligne, consulté le )
- MEROPS
- Jian Yang, « TrED : the Trichophyton rubrum Expression Database », BMC Genomics, vol. 8, no 1, , p. 250 (ISSN 1471-2164, DOI 10.1186/1471-2164-8-250, lire en ligne, consulté le )
- Sandy Vermout, Contribution à l’étude fonctionnelle des dipeptidyl peptidases et de la métalloprotéase MEP3 sécrétées par Microsporum canis, , thèse, Université de Liège
- Michel Monod, « Aminopeptidases and dipeptidyl-peptidases secreted by the dermatophyte Trichophyton rubrum », Microbiology, Reading, England, vol. 151, no Pt 1, , p. 145-155 (ISSN 1350-0872, DOI 10.1099/mic.0.27484-0)
- Fernanda C. A. Maranhão, « Membrane transporter proteins are involved in Trichophyton rubrum pathogenesis », Journal of Medical Microbiology, vol. 58, no 2, , p. 163-168 (ISSN 0022-2615 et 1473-5644, DOI 10.1099/jmm.0.002907-0, lire en ligne, consulté le )
- Nilce M. Martinez-Rossi, « Role of pH in the pathogenesis of dermatophytoses », Mycoses, vol. 55, no 5, , p. 381–387 (ISSN 1439-0507, DOI 10.1111/j.1439-0507.2011.02162.x, lire en ligne, consulté le )
- Nalu T.A. Peres, « Transcriptional profiling reveals the expression of novel genes in response to various stimuli in the human dermatophyte Trichophyton rubrum », BMC Microbiology, vol. 10, no 1, , p. 39 (ISSN 1471-2180, DOI 10.1186/1471-2180-10-39, lire en ligne, consulté le )
- Nalu Teixera de Aguiar Peres, « Dermatophytes: host-pathogen interaction and antifungal resistance », Anais brasileiros de dermatologia, vol. 85, no 5, , p. 657-667 (ISSN 1806-4841)
- R.C. Summerbell, « rRNA Gene Internal Transcribed Spacer 1 and 2 Sequences of Asexual, Anthropophilic Dermatophytes Related to Trichophyton rubrum », J Clin Microbiol, vol. 37, no 12, , p. 4005-4011 (ISSN 0095-1137, PMID 10565922, DOI 10.1128/JCM.37.12.4005-4011.1999)
- Claus Seebacher, Jean-Philippe Bouchara, Bernard Mignon, « Updates on the epidemiology of dermatophyte infections », Mycopathologia, vol. 166, nos 5-6, , p. 335-352 (ISSN 0301-486X, DOI 10.1007/s11046-008-9100-9)
- J.W. Rippon, « The changing epidemiology and emerging patterns of dermatophyte species », Current topics in medical mycology, vol. 1, , p. 208-234 (ISSN 0177-4204)
Liens externes
- (en) Référence Catalogue of Life : Trichophyton rubrum (Castell.) Sabour. (consulté le )
- (en) Référence Index Fungorum : Trichophyton rubrum (+ MycoBank)
- (en) Référence MycoBank : Trichophyton rubrum