Trichophyton
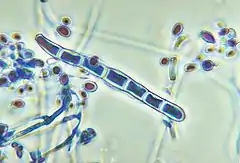
Trichophyton est un genre de champignons responsable de dermatophytose (mycose de la peau), caractérisée par le développement de microconidies et macroconidies, à parois lisses. Ces champignons sont capables d'infecter autant des animaux que des humains, Trichophyton rubrum étant l'espèce la plus répandue.
Étymologie
Le nom de genre Trichophyton est un terme de latin scientifique composé à partir de deux étymons grecs θρίξ thríx, « cheveux » et de φυτόν phutón, « plante »[1].
Cycle de vie
Les différentes espèces de Trichophyton peuvent accomplir leur cycle de vie en faisant intervenir la reproduction et la multiplication asexuée, mais seule la multiplication asexuée a lieu in vivo.
Les hyphes produisent un conidiophore qui libère des spores qui pourront ensuite germer pour à leur tour produire des hyphes. Deux types de spores peuvent être produites: unicellulaires (microconidies) ou pluricellulaires (macroconidies).
Mécanisme d'infection
L'infection se déroule en trois étapes. Le point de départ est le contact entre une spore et l'épiderme ou tout autre partie du corps contenant de la kératine. Par un mécanisme complexe d'interactions entre des molécules de surface et des protéases, la spore adhère à l'épiderme. La spore agit alors comme un capteur pour déterminer les conditions favorables de croissance du champignon. Lorsque les conditions sont réunies, la spore est réhydratée ce qui permet la formation de nouveaux composants de la paroi cellulaire et une réactivation du métabolisme. Des acides nucléiques et des protéines sont produits en même temps, permettant la création de nouvelles structures tubulaires, de composants cellulaires mais aussi de renforcer l'hyphe. Lorsque la cellule est assez développée, elle va se diviser pour former plusieurs cellules interconnectées par un pore formé par un septum. Ce procédé est appelé la germination et a lieu en continu permettant ainsi la croissance de l'hyphe à partir de chaque compartiment cellulaire. La dernière étape est l'invasion de l'épiderme par l'hyphe. Au cours de l'invasion, l'hyphe crée des ramifications et des colonies dans les différentes couches de l'épiderme et dégrade la kératine, résultant en la déstructuration de l'épiderme. Du fait de la perte d'intégrité de l'épiderme, l'expression des kératinocytes est augmentée afin de lutter contre l'invasion. Le système immunitaire est alors déclenché avec la libération de cytokines et de peptides antimicrobiens[2].
Diagnostic
Le diagnostic repose sur quatre étapes : l'interrogatoire, le prélèvement, l'examen direct et la culture afin d'identifier la souche et choisir le traitement approprié. L'examen direct permet de visualiser les éléments fongiques se trouvant au niveau de l'infection. Il permet un examen rapide pour apporter un traitement provisoire pendant le temps de culture du champignon.
Traitements
Traitements actuels[3]
A la suite du diagnostic, le médecin prescrit un traitement qui prend en compte le dossier médical du patient (maladies chroniques, autres prescriptions), afin d'éviter d'interférer avec tout autre traitement, la vitesse d'évolution et la date d'apparition des lésions ainsi que la présence d'autres symptômes. Les traitements peuvent se trouver sous forme orale ou cutanée et sont divisés en différentes catégories en fonction de leur cible. De nos jours, il existe de nombreuses molécules permettant d'inhiber la synthèse de composant cellulaires, de bloquer la réplication du champignon, la synthèse d'ADN ou d'ARN, ou encore interagir avec les différentes organelles cellulaires ou la membrane.
Les traitements sont longs et doivent être continués même après la disparition des symptômes pour permettre l'élimination de toutes les spores. Un arrêt prématuré du traitement conduira à une reprise de l'infection.
Nouveaux traitements
Face à l'émergence de souches résistantes, les chercheurs tentent d'identifier de nouvelles molécules permettant d'éliminer l'infection. Cependant, du fait de procédés biologiques partagés entre les champignons et les humains, il est difficile de développer un médicament permettant à la fois d'inhiber la croissance du champignon sans endommager les cellules hôtes. Ces dernières années, les produits à base de plantes ont révélé des résultats promettant[3].
Résistances aux traitements
De nos jours, des souches résistantes ont été identifiées, et certains mécanismes de résistance élucidés. Si les caractéristiques structurales du champignon sont un de ses atouts pour résister aux traitements, il peut aussi modifier ses mécanismes de signalisation en réponse au stress induit par le traitement. Tous ces mécanismes ont pour objectif d'empêcher l'action du traitement, que ce soit en empêchant la fixation sur la molécule cible, ou alors en éliminant l'agent toxique de la cellule[4].
Détoxification cellulaire
Les cellules du champignons sont capables de produire un grand nombre d'enzymes en fonction de leur environnement, dont certaines sont capables d'éliminer le pouvoir cytotoxique du traitement. Par ailleurs, une surproduction de catalases peut être observée, qui permettent de protéger les cellules de l'hyphe des effets apoptotiques induits par le traitement.
Efflux cellulaire
Les cellules de l'hyphe comportent des transporteurs cellulaires sur leur membrane appelés pompes efflux. Ce sont des protéines membranaires capables de se fixer à différents composants chimiques pour les extraire de la cellule. Dans le cas d'un traitement antifongique, le champignon est capable de surexprimer les gènes codant ces pompes, ce qui lui permet d'empêcher l'accumulation du traitement dans la cellule.
Protéines de choc thermique
Les protéines de choc thermique sont une famille de chaperonnes impliquées dans la traduction, la transcription, les modifications post-transcriptionnelles, le repliement, l'agrégation et désagrégation des protéines. Lorsque la cellule fongique est soumise à un stress, notamment du fait d'un traitement, ces protéines vont être surexprimées afin d'aider la cellule à s'adapter à ce stress. C'est un mécanisme indirect de résistance aux traitements.
Modification transcriptionnelles des protéines kinases
Les protéines kinases impliquées dans la transmission des stimuli cellulaires peuvent être surexprimées ou sousexprimées afin de permettre à la cellule d'échapper à l'agent antifongique.
Mutations dans la séquence protéique des cibles des traitements
Au-delà de modification d'expression de protéines, des mutations ponctuelles sont apparues dans des gènes permettant la synthèse de composants cellulaires. C'est notamment le cas pour le gène codant l'enzyme permettant la synthèse de l'ergostérol composant la membrane cellulaire. Ces mutations résultent en une substitution d'un acide aminé dans la région où la molécule antifongique se fixe pour empêcher la synthèse d'ergostérol. Cette substitution conduit à l'échec du traitement mais n'empêche pas une correcte production d'ergostérol[5] - [6] - [7].
Facteurs favorisant l'infection
De nombreux facteurs ont été identifiés comme augmentant le risque d'infection. Ils sont le résultat d'interactions entre le pathogène et l'hôte. Du côté de l'hôte, l'état de santé, le mode de vie et des prédispositions génétiques peuvent le rendre plus sensible à une infection. De nombreux sports ont notamment été pointés du doigt comme augmentant les risques, et des recommandations sanitaires ont été développées afin de lutter contre ces infections. Du côté du pathogène, l'environnement, les types d'enzymes sécrétées et son génome peuvent faciliter l'adhésion au moment de l'infection, ou sa résistance aux traitements[8].
Espèces et leur milieu naturel
- Trichophyton ajelloi, géophile
- Trichophyton concentricum, anthropophile
- Trichophyton equinum, zoophile (cheval)
- Trichophyton flavescens, géophile (plumes)
- Trichophyton gloriae, géophilie
- Trichophyton indotineae
- Trichophyton megnini, anthropophile
- Trichophyton mentagrophytes
- Trichophyton mentagrophytes var. erinacei, zoophile (hérisson)
- Trichophyton mentagrophytes var. interdigital, anthropophile
- Trichophyton phaseoliforme, géophile
- Trichophyton rubrum, anthropophile
- Trichophyton rubrum downy strain, anthropophile
- Trichophyton rubrum granular strain, anthropophile
- Trichophyton schoenleinii, anthropophile
- Trichophyton simii, zoophile (singe)
- Trichophyton soudanense, anthropophile
- Trichophyton terrestre, géophile
- Trichophyton tonsurans, anthropophile
- Trichophyton vanbreuseghemii, géophile
- Trichophyton verrucosum, zoophile (bovins, cheval)
- Trichophyton violaceum, anthropophile
- Trichophyton yaoundei, anthropophile
Références
- voir trichophyton
- (en) Émilie Faway, Catherine Lambert de Rouvroit et Yves Poumay, « In vitro models of dermatophyte infection to investigate epidermal barrier alterations », Experimental Dermatology, vol. 27, no 8, , p. 915–922 (ISSN 1600-0625, DOI 10.1111/exd.13726, lire en ligne, consulté le )
- (en) S. Gnat et D. Łagowski, « Major challenges and perspectives in the diagnostics and treatment of dermatophyte infections », sur Journal of applied microbiology, (PMID 32048417, consulté le ).
- (en) Nilce M. Martinez-Rossi, Tamires A. Bitencourt, Nalu T. A. Peres et Elza A. S. Lang, « Dermatophyte Resistance to Antifungal Drugs: Mechanisms and Prospectus », Frontiers in Microbiology, vol. 9, (ISSN 1664-302X, DOI 10.3389/fmicb.2018.01108, lire en ligne, consulté le )
- (en) Yamada T et Maeda M, « Terbinafine Resistance of Trichophyton Clinical Isolates Caused by Specific Point Mutations in the Squalene Epoxidase Gene », sur Antimicrobial agents and chemotherapy, (PMID 28416557, consulté le )
- (en) Rudramurthy Sm et Shankarnarayan Sa, « Mutation in the Squalene Epoxidase Gene of Trichophyton interdigitale and Trichophyton rubrum Associated with Allylamine Resistance », sur Antimicrobial agents and chemotherapy, (PMID 29530857, consulté le )
- (en) Dominik Łagowski, Sebastian Gnat, Aneta Nowakiewicz et Marcelina Osińska, « Intrinsic resistance to terbinafine among human and animal isolates of Trichophyton mentagrophytes related to amino acid substitution in the squalene epoxidase », Infection, vol. 48, no 6, , p. 889–897 (ISSN 1439-0973, PMID 32770418, PMCID PMC7674369, DOI 10.1007/s15010-020-01498-1, lire en ligne, consulté le )
- (en) Gnat S et Nowakiewicz A, « Host- and pathogen-dependent susceptibility and predisposition to dermatophytosis », sur Journal of medical microbiology, 2019 jun (PMID 31050630, consulté le )
- (en) Référence Index Fungorum : Trichophyton (+ liste espèces) (+ MycoBank)
- (en) Référence Catalogue of Life : Trichophyton (consulté le )
- (en) Référence NCBI : Trichophyton (taxons inclus)
- https://web.archive.org/web/20070524031748/http://www.mycology.adelaide.edu.au/Fungal_Descriptions/Dermatophytes/Trichophyton/
- (en) Faway, É., Rouvroit, C.L. de, and Poumay, Y. (2018) In vitro models of dermatophyte infection to investigate epidermal barrier alterations. Experimental Dermatology 27: 915–922. https://onlinelibrary.wiley.com/doi/full/10.1111/exd.13726
- (en) Gnat, S., Łagowski, D., and Nowakiewicz, A. (2020) Major challenges and perspectives in the diagnostics and treatment of dermatophyte infections. Journal of Applied Microbiology 129: 212–232. https://pubmed.ncbi.nlm.nih.gov/32048417/
- (en) Gnat, S., Nowakiewicz, A., Łagowski, D., and Zięba, P. (2019) Host- and pathogen-dependent susceptibility and predisposition to dermatophytosis. Journal of Medical Microbiology, 68: 823–836. https://pubmed.ncbi.nlm.nih.gov/31050630/
- (en) Łagowski, D., Gnat, S., Nowakiewicz, A., Osińska, M., and Dyląg, M. (2020) Intrinsic resistance to terbinafine among human and animal isolates of Trichophyton mentagrophytes related to amino acid substitution in the squalene epoxidase. Infection https://doi.org/10.1007/s15010-020-01498-1. Accessed November 12, 2020. https://link.springer.com/article/10.1007/s15010-020-01498-1
- (en) Martinez-Rossi, N.M., Bitencourt, T.A., Peres, N.T.A., Lang, E.A.S., Gomes, E.V., Quaresemin, N.R., et al. (2018) Dermatophyte Resistance to Antifungal Drugs: Mechanisms and Prospectus. Front Microbiol 9 https://www.frontiersin.org/articles/10.3389/fmicb.2018.01108/full. Accessed November 12, 2020. https://www.frontiersin.org/articles/10.3389/fmicb.2018.01108/full
- (en) Monod, M., Feuermann, M., Salamin, K., Fratti, M., Makino, M., Alshahni, M.M., et al. (2019) Trichophyton rubrum Azole Resistance Mediated by a New ABC Transporter, TruMDR3. Antimicrobial Agents and Chemotherapy 63: 19. https://aac.asm.org/content/63/11/e00863-19
- (en) Rudramurthy, S.M., Shankarnarayan, S.A., Dogra, S., Shaw, D., Mushtaq, K., Paul, R.A., et al. (2018) Mutation in the Squalene Epoxidase Gene of Trichophyton interdigitale and Trichophyton rubrum Associated with Allylamine Resistance. Antimicrob Agents Chemother 62, /aac/62/5/e02522-17.atom. https://pubmed.ncbi.nlm.nih.gov/29530857/
- (en) Yamada, T., Maeda, M., Alshahni, M.M., Tanaka, R., Yaguchi, T., Bontems, O., et al. (2017) Terbinafine Resistance of Trichophyton Clinical Isolates Caused by Specific Point Mutations in the Squalene Epoxidase Gene. Antimicrob Agents Chemother 61, https://pubmed.ncbi.nlm.nih.gov/28416557/