Système de polarité segmentaire
Le système de polarité segmentaire est régulé par l'expression de divers gènes qui permettent de configurer les patrons de la segmentation qui est la répétition d’une unité structurée composé d’une série de caractères pouvant former un corps unique, le long de l’axe antéro-postérieur. Ces gènes délimitent les frontières des parasegments du blastoderme qui est une couche cellulaire primitive de l'embryon, grâce à l’induction de facteur de transcription, aux protéines sécrétées et de ses récepteurs pour la communication entre cellules adjacentes des frontières. Les groupes ayant un modèle de segmentation sont les annélides, les arthropodes et les chordés. Ce système de polarité a été détaillé à l’aide d’un modèle, Drosophila melanogaster.
Origine et évolution de la segmentation
La segmentation se retrouve dans trois embranchements, les annélides, les arthropodes, et les chordés. L’évolution de la segmentation a tout de même été utile pour créer des relations avec le système métazoaire, les bilatérales. À la fin du XIXe siècle, la découverte d’un segment d’un ancêtre commun regroupant ces trois embranchements suppose que la segmentation est un caractère homologue qui partage le même système de développement. Cependant, à la fin du XXe siècle, avec la distinction entre les protostomiens et les deutérostomiens et l’affinité proche de certains groupes des bilatéraux et des chordés, trois autres hypothèses ont été émis. La première hypothèse est que chaque embranchement a acquis la segmentation indépendamment. La deuxième hypothèse est que la segmentation est apparue indépendamment chez les chordés dans laquelle les caractères de segmentation sont similaires entre les annélides et les arthropodes. Enfin, la troisième hypothèse est que la segmentation est homologue lors de l’évolution des bilatéraux. Ces deux dernières hypothèses se basent sur des données moléculaires dans le développement d’un segment de l’ancêtre commun durant l’embryogenèse[1].
Chez les arthropodes
Ce système de polarité segmentaire, contrôlé par Wingless et Engrailed et son interaction cellulaire sont conservés entre les groupes des arthropodes au cours de l’évolution. En effet, Drosophila et Tribolium possèdent le réseau de polarité segmentaire, ainsi que le système segmentaire avec les gènes Pair-Rules. Ce mécanisme est activé et est nécessaire dans le blastoderme de l’insecte durant l’embryogenèse afin de prédisposer chaque segment. La formation des segments est différente selon le temps de développement pour les insectes. Par exemple, chez les insectes qui germent de courte durée, comme Tribolium, les segments de la tête sont formés dans le blastoderme et les autres segments sont créés progressivement avec la croissance du postérieur. Chez les insectes qui germent à un temps plus long, comme Drosophila, tous les segments sont configurés dans le blastoderme[1].
Activation des gènes de polarité segmentaire

Le modèle permettant de comprendre au mieux le développement de l’embryon d’un insecte est Drosophila melanogaster. Trois types de gènes sont importants pour la formation de segments tels que : les gènes maternels, les gènes de segmentation, les gènes homéotiques. Les gènes de segmentation sont divisés en trois catégories, les gènes Gap, les gènes Pair-Rules (en) et les gènes de polarité segmentaire. Les gènes Gap permettent de configurer l’axe antéro-postérieur en fonction de leur gradient de concentration des ARNm[2].
Suite à l’expression des gènes comme Hunchback, Bicoid et Nodal (en) formant cet axe[2], les gènes Pair-Rule dont Even-Skipped et Fushi Tarazu, configurent la segmentation en 7 bandes parasegmentaires. Avant le système de polarité segmentaire, lorsque l’embryon est divisé en 14 bandes cellulaires le long de l’axe antéro-postérieur, Even-Skipped et Fushi Tarazu contrôlent la mise en place l’expression d’Engrailed dans les bandes cellulaires impaires avec ses séquences cis-régulation. L'augmentation de leur expression dans les cellules situées antérieurement aux parasegments activent l'expression d'Engrailed et donc activent le réseau de polarité segmentaire. À cette période, Wingless et Engrailed apparaissent dans les bandes cellulaires localisées le plus antérieur et le plus postérieur de chaque parasegment.
Les mutants de Fushi Tarazu démontrent l’absence des bandes formées par Engrailed et l’absence de formation de la moitié des parasegments. Parallèlement avec les mutants d’Even-Skipped, les bandes cellulaires d’Engrailed et ses parasegments ne sont pas formées. De plus, l’absence de ce dernier montre qu’il y a une absence de segmentation et qu’Even-Skipped est nécessaire et important pour les gènes de polarité segmentaire. En effet, en présence d’Even-Skipped, toutes les bandes paires sont exprimées par Engrailed avec les domaines FTZ et peu de bandes impaires sont exprimées. En opposé, Wingless qui est une protéine sécrétée pour la communication cellulaire[3], est réprimée par les gènes de Pair Rules dans les cellules se trouvant à l’extrémité postérieure d’un parasegment. Il est donc exprimé dans les cellules postérieures situées au plus proche du front du parasegment, à côté des cellules exprimant Engrailed pour former les cellules antérieures du parasegment[4]. Ainsi, Engrailed, issu d'un facteur de transcription du noyau[5], est exprimé dans les cellules antérieures pour former les cellules postérieures d’un parasegment donnant lieu la partie antérieure d’un segment. Le contrôle des gènes Pair-Rules est un mode de régulation du système de polarité segmentaire[4].
Résumé du système de polarité segmentaire
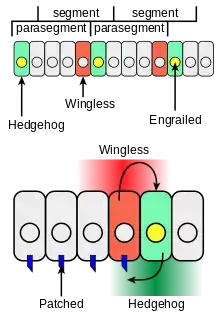
La communication cellulaire produite par Wingless (Wg) et Engrailed (En) impliquent aussi l’induction de récepteurs. de protéines sécrétées et accessoires par leurs facteurs de transcription pour délimiter le patron des segments avec l’aide d’un ensemble de gènes comme Hedgehog, Patched, Frizzled. Au cours de la gastrulation, Wingless et Engrailed deviennent dépendantes de leur expression[6]. Ce système permet de réguler l'expression de ces gènes et de maintenir le réseau de polarité segmentaire pour la différenciation des cellules formant le front d'un parasegment jusqu'à l'arrivée des gènes homéotiques.
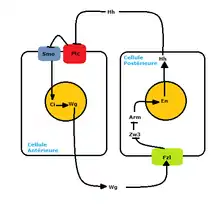
Une fois que les gènes Pair Rule ont assuré l'activation de ce réseau de polarité segmentaire, il y a l'expression de Hedgehog activé par le facteur de transcription d'Engrailed. En effet, les cellules antérieures du parasegment exprimant Engrailed expriment elles-aussi Hedgehog. Ce dernier est considéré comme un gène de polarité segmentaire car son rôle est de signaler aux cellules voisines qu'elles seront les cellules postérieures du parasegment. Le récepteur de ce signal, Patched, se situe à la membrane des cellules postérieures des parasegments[6]. Il permet de maintenir le réseau en activant par la suite l'expression du gène Wingless qui assure la production de son signal. Lors de l’émission du signal Wingless, il se fixe alors à son récepteur situé à la membrane de la cellule antérieure qui est Frizzled, une protéine à 7 passages membranaires. Cette boucle de rétroaction positive permet de fixer les parasegments tout le long de l’embryogénèse[7].
Expression/Répression du signal Wingless
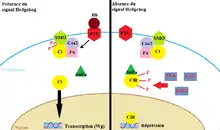
Lorsqu'il y a l'absence du signal Hedgehog, la cellule postérieure ne produit pas le facteur de transcription de Wingless vu que le réseau de polarité segmentaire a une organisation de boucle de rétroaction positive. En effet, en absence d'Hedgehog, son récepteur Patched inhibe l'activité d'une autre protéine transmembranaire qui est Smoothened et qui forme un grand complexe contenant d'autres protéines telles que : Cubitus interruptus (Ci), Répresseur de Fused (Su(Fu)), Fused (Fu), Costal-2 (Cos2). Son inhibition inhibe ainsi la translocation de Cubitus interruptus du cytoplasme vers le noyau. Son empêchement mène ainsi à la répression de l'expression de Wingless dans le noyau sachant que Cubitus interruptus est son facteur de transcription. Cet empêchement est aussi amplifié car trois kinases différentes, PKA (protéine kinase A), GSK (glycogène synthase kinase 3), CK1 (caséine kinase 1) le phosphorylent en rendant Cubitus interruptus comme un répresseur de Wingless (CiR) en inhibant sa transcription dans le noyau. De plus, le répresseur de Fused empêche l'activité de Cubitus interruptus.
Lorsqu'il y a la présence du signal Hedgehog, sa liaison avec Patched active le complexe Smoothened et ses protéines. Cette activation arrête la production de CiR et Cubitus interruptus se déplace librement dans le noyau afin d'activer les gènes cibles du signal Hedgehog, dont Wingless[8]. D'ailleurs, chez la drosophile, Cubitus interruptus (ci), protéine se situant dans les cellules antérieures des parasegments, a un rôle essentiel dans la régulation de la voie de signalisation de Wingless et d’Hedgehog. Ce dernier est un gène de polarité segmentaire qui permet de maintenir l’expression du gène Wingless en encodant une protéine de type doigt de zinc. Il a aussi pour rôle de spécialiser la cuticule nue de chaque segment épidermique. Il est régulé de façon post-transcriptionnelle lorsque l’embryon est composé de 11 bandes cellulaires et son gradient d’ARNm est plus élevé au niveau des fronts des parasegments grâce au signal de Hedgehog. De plus, chez les mutants Fused, qui est une protéine sérine/thréonine kinase, Hedgehog et Wingless démontrent qu’il y a une absence de la distribution de ce gène dans les cellules antérieures. Les activités des gènes Fused et Cubitus interruptus sont indirectement aussi régulées par le récepteur Patched de Hedgehog. Le gène ci requiert l’activité kinase de Fused sinon son activité est instable. Au stade 11, la transcription de la protéine Patched est dépendante de la transcription de Cubitus interruptus et agit de manière opposée de ce dernier sur l’expression de Wingless en l’inhibant[3].
Expression/Répression du signal Hedgehog
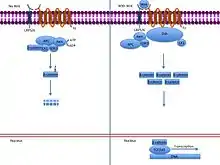
En parallèle, dans les cellules exprimant le signal Hedgehog par le facteur de transcription Engrailed, il y a de même une autre cascade de signalisation de Wingless avec l'activation d'un complexe de protéines appelées Dishevelled, Zeste-white3 et Amardillo considérées comme des gènes de polarité segmentaire chez la Drosophile[9].
Lorsqu'il y a l'absence du signal Wingless et donc à l'absence d'une liaison entre Wingless et Frizzled, Dishevelled, une protéine semi-membranaire et multifonctionnelle, est inactivée. Son inactivation mène à l'activation de la protéine kinase Zeste-White-3 qui a pour but de phosphoryler Armadillo, une β-caténine. La phosphorylation de cette dernière mène à sa dégradation et ainsi à la répression de la transcription du gène Engrailed.
Or, lorsqu'il y a l'émission du signal Wingless, Frizzled le perçoit et active Dishevelled grâce à sa liaison PDZ. Par la suite, Dishevelled en étant phosphorylé inhibe la protéine kinase Zeste-White3 qui encode un glycogène synthase et kinase 3, GSK3. Son inactivation mène à la non-phosphorylation de la β-caténine, soit Armadillo. La libération d'Armadillo peut traverser le noyau et se lier au facteur de transcription. Cette liaison permet ainsi l’activation de l’expression de Hedgehog. Cette boucle de rétroaction implique alors la réception du signal Hedgehog par Patched et la réception du signal Wingless par Frizzled. Ce mode d’auto-régulation de Wingless est une des régulations de la voie de signalisation du système de polarité segmentaire[9].
Par ailleurs, une étude a été faite pour observer de plus près la signalisation de Wingless dans le développement de l'embryon de la Drosophile. Pour débuter, les 14 segments divisés de l'embryon ont été facilement démarqués par l'alternance de deux zones différentes dont des zones de cuticule nue et de zones ayant une excroissance cuticulaire sur les cellules épidermiques qui sont démarquées par les gènes de polarité segmentaire, en outre le gène Wingless. En effet, des observations ont été effectuées chez des embryons homozygotes et mutants wg et leurs résultats montrent qu'il y avait une disparition des zones de cuticules nues et une perte de polarité des autres zones ayant une excroissance cuticulaire. Ces résultats ont montré que le gène Wingless avait un rôle majeur sur la distinction de ces deux zones et que l'expression d'Engrailed est dépendante de sa présence dans les cellules antérieures de chaque segment. De plus, des expériences plus développées ont été effectuées sur d'autres gènes comme Zw3 pour voir son rôle dans le développement de l’embryogenèse. Sachant que Zw3 est une kinase qui inhibe la transcription du gène Engrailed, lorsqu'on le mute, il y a l'activation d'Engrailed et morphologiquement, l'embryon de ses mutants est recouvert de zones de cuticule nue[9].
Dishevelled
La nécessité de tous ces gènes permet le bon fonctionnement de la polarité segmentaire de l’embryon de la Drosophile. Ce gène est requis pour l’établissement des cellules autonomes et la formation des segments. En effet, Dishevelled encode une protéine intracellulaire qui partage un motif d’acides aminés avec d’autres protéines pour répondre au signal de Wingless qui est perçu par le récepteur Frizzled. Une fois que le signal est perçu par Dishevelled, il permet d'activer l'expression de la protéine Armadillo qui transmet la perception du signal des cellules épithéliales adjacentes et leur cytosquelette en se relâchant dans le cytoplasme. Une mutation sur ce gène arrête la fonction d'Armadillo. De plus, des expériences démontrent que lorsqu'il y a des mutations sur les cellules exprimant Wingless, il y a une baisse d'activité ou perte d'activité de Dishevelled[10].
Armadillo
Armadillo (arm), considéré comme gène de polarité segmentaire étant distribué dans les cellules contenant Wingless, est aussi nécessaire pour réglementer le réseau de polarité segmentaire car l’expression de son ARNm est dépendant du signal de Wingless tout comme Zeste-White3/Shaggy (zw3)[11]. D'ailleurs, lorsqu'il n'y a pas de signal, ces protéines sont stockés dans le cytoplasme en se liant à des molécules α-caténines au domaine cytoplasmique des cadhérines[9]. De plus, des modifications du gène proche de la membrane plasmique montrent qu’il est nécessaire pour les cellules cibles du signal Wingless. Chez les mutants Wg, il n’y a pas d’accumulation d’Armadillo. Son interaction avec Wingless est importante pour cibler les cellules formant la partie antérieure du segment et mène à des phénotypes similaires[11].
Cubitus interruptus
Cubitus interruptus (ci), faisant partie des gènes de polarité segmentaire et dépendant du signal Hedgehog, est essentiel au fonctionnement du système car les mutations de ce gène affectent l’embryogenèse. Ce gène est issu d'un facteur de transcription du noyau de la cellule[5]. Des manipulations sur les mutations de ce gène ont été effectuées et ont démontré qu'il y a trois types de mutations qui modifient la fonctionnalité de Cubitus interruptus. En effet, la première classe de mutation arrête le signal de Wingless dans les régions postérieures de deux parasegmentaires de la drosophile. Le deuxième genre de mutations sur ce gène provoque l’élimination de celui-ci au début de l’embryogenèse et mène à la délétion des cellules exprimant ce gène jusqu’aux cellules exprimant Engrailed. Enfin, le dernier type de mutations élimine Cubitus interruptus plus tard dans le développement de l’embryon et interrompt la formation du cuticule ventrale. D’ailleurs, ces deux derniers classes de mutations engendrent la répression de Wingless et de son signal[3].
Gooseberry
Gooseberry (gsb) est un autre gène de polarité segmentaire qui encode un facteur de transcription contenant un domaine pair et un homéodomaine permet de maintenir l’expression de Wg à travers une boucle de rétroaction entre Gsb et Wg lors du développement de l’embryon chez la Drosophile[12]. Son rôle est d'emplacer les segments en fonction de la polarité (antérieur vers postérieur)[13]. Lorsqu’on mute ce gène dans les embryons, la partie postérieure et nue de chaque segment est remplacé par le motif antérieur. En effet, l’activation de la région cis-régulatrice de gsb est responsable pour la continuité de l’expression de Gooseberry par Wingless par communication paracrine et autocrine. D’après les expériences, Gooseberry agit comme un répresseur pour la formation des denticules ventraux durant l’embryogenèse. En effet, Engrailed n’agit pas comme répresseur de la formation des segments sachant qu’il y a la production de denticules dans les cellules antérieures. Ainsi, le signal de Wingless agit de façon directe pour réprimer la formation des denticules grâce à deux autres gènes de polarité segmentaire tels que Dishevelled et Armadillo de manière autonome. Le signal de la protéine Wingless agit de même de façon indirecte en activant Gooseberry[12].
Gènes de polarité segmentaire pour la neurogénèse
Certains gènes de polarité segmentaires sont requis pour la détermination et l’identification des neurones lors du développement du système nerveux central dans l’embryon de la drosophile. Pour les embryons n'exprimant pas ces gènes suivant : Fused, Armadillo et Cubitus interruptus, le système nerveux est « normal ». Cependant, lorsqu’il y a des mutations sur les autres gènes clés de la polarité segmentaire, il y a des délétions et des modifications structurales dans l’épiderme de l’espèce le long de l’axe antéro-postérieur. Ces modifications interromptent ainsi la spécialisation des cellules en neurones. En effet, chez les mutants Hedgehog de l’embryon, il y a une désorganisation du système nerveux central durant la fin du cycle de la neurogénèse. Les mutations sur Wingless, Patched et Gooseberry engendrent des défauts spécifiés dans le système nerveux central. Des mutants Patched montrent que ce gène est important pour la formation d’un groupe de neuroblastes et de la neurogénèse. La délétion des cellules est remplacée par une duplication de cellules venant de la partie postérieure du segment. Cette modification spatiale duplique les neurones et les neuroblastes exprimés par Engrailed. Cette modification change la région postérieure de chaque segment du système nerveux central. Les mutants Wingless démontrent une malformation au niveau phénotypique ce qui engendre une mauvaise communication entre deux neurones et ces cellules ne pourront pas exprimer Even-Skipped[14].
D'ailleurs, Gooseberry est un gène comportant un locus important pour la neurogénèse, appelé Gooseberry neuro (GSN) qui code un facteur de transcription qui se lie à l'ADN conservé dans le noyau. GSN est exprimé dans les cellules composant le système nerveux central et le tube neural, c'est-à-dire les cellules épidermales qui expriment Wingless au milieu de chaque segment[13]. Les mutations sur Gooseberry ont un impact sur la croissance des cellules neuronales en altérant la capacité de s’identifier dans les régions épidermiques[14].
Chez les annélides
Signalisation d'Engrailed et Wingless
En prenant le modèle Platynereis dumerilii, il y a le même système de polarité segmentaire avec les gènes Engrailed et Wingless retrouvé chez les Arthropodes. Ces deux gènes sont exprimés dans les bandes ectodermiques à la frontière de chaque segment avant, durant et après la formation des segments et sont importantes pour cette dernière. Avec cette similitude retrouvée chez les arthropodes, ce système est sûrement dû à une convergence évolutionnaire ou même un système homologue d’un ancêtre commun regroupant les deux embranchements. D’après des manipulations avec l’expression des gènes Pdu-en et Pdu-wg, on émet l’hypothèse de l’évolution de ces gènes communs venant d’un ancêtre commun, Urpotostome. Cette unité segmentaire ancestrale régulée par le système de polarité segmentaire correspond aux parasegments chez les arthropodes et aux segments adultes chez les annélides[15]. Cependant, pour d’autres espèces d’annélides, la fonction générale d’Engrailed pour la segmentation est perdue et seulement exprimée dans le nerf médullaire. Or ceci peut-être aussi expliqué par la perte de fonction d’Even-Skipped chez quelques espèces d'arthropodes. Cette perte peut aussi démontrer la convergence évolutionnaire des annélides et des arthropodes[16].
Signal Hedgehog
Plus de recherches ont été effectuées sur Platynereis dumerilii avec la signalisation Hedgehog et son récepteur Patched. Dans ce cas, Hedgehog et son récepteur agissent comme des gènes régulant la formation des segments et non de l’établissement de ces derniers. Cette signalisation commune aux arthropodes et aux annélides démontre qu’il y a une origine commune dans la segmentation chez les protostomes. Premièrement, la segmentation de Platynereis utilise le système de polarité segmentaire avec Engrailed, pour la partie antérieure du segment et Wingless pour la partie postérieure du segment. Deuxièmement, nous avons l’expression de Cubitus interruptus dans les cellules les plus lointaines des fronts de segments aux fronts des segments. Les cellules les plus lointaines expriment le plus ce gène et son gradient diminue lorsqu’on s’approche des limites du segment. Puis, Patched, est plus exprimé à chaque extrémité de chaque segment[16]. Cependant, l’action du signal d’Hedgehog sur les parties antérieures n’a pas encore été prouvée[16].
Voir aussi
Articles connexes
Références
- (en) Gregory K. Davis and Nipam H. Patel, « The origin and evolution of segmentation », Trends in genetics,
- (en) Gurbachan S. Miglani, Developmental Genetics, I. K. International Pvt Ltd, , 780 p. (ISBN 978-81-88237-59-3, lire en ligne)
- (en) Diane C. Slusarski, Cynthia Kelsey Motzny and Robert Holmgren, « Mutations that Alter the Timing and Pattern of cubitus interruptus Gene Expression in Drosophila melanogush », Genetics, , p. 139: 229-240
- (en) Miki Fujioka, James B. Jaynes, and Tadaatsu Goto, « Early even-skipped stripes act as morphogenetic gradients at the single cell level to establish engrailed expression », Development., , p. 121(12): 4371–4382 (lire en ligne)
- (en) Maxine Singer et Paul Berg, Exploring Genetic Mechanisms, Californie, University Science Books, p. 487
- (en) Tetsuya Tabata and Thomas B. Kornberg, « Hedgehog Is a Signaling Protein with a Key Role in Patterning Drosophila lmaginal Discs », Cell, , Vol 70, 89-102
- (en) Michael M. Cox, Molecular Biology, Palgrave Macmillan, , 750 p. (ISBN 978-1-4641-6292-3, lire en ligne)
- (en) Lewis Wolpert,Cheryll Tickle,Alfonso Martinez Arias, Principles of Development, Oxford University Press, , 82 p. (lire en ligne), p. 81-83
- (en) Yohanns Bellaïche et Norbert Perrimon, « La voie de signalisation Wingless chez la drosophile », médecine/sciences, , p. 13 : 166-74 (lire en ligne)
- (en) Heidi Theisen, Judith Purcell, Mary Bennett, Dipul Kansagara, Adeela Syed and J. Lawrence Marsh, « Dishevelled is required during wingless signaling to establish both cell polarity and cell identity », Development, , p. 347-360
- (en) Bob Riggleman, Paul Schedl, and Eric Wieschaus, « Spatial Expression of the Drosophila Segment Polarity Gene armadillo Is Posttranscriptionally Regulated by wingless », Cell Press, , Vol. 63, 549-560
- (en) Xuelin Li and Markus Noll, « Role of the gooseberry gene in Drosophila embryos: maintenance of wingless expression by a wingless gooseberry autoregulatory loop », The EMBO Journal, , vol. 12 no. 12 pp.4499 - 4509 (lire en ligne)
- (en) Haihuai He, Markus Noll, « Differential and redundant functions of gooseberry and gooseberry neuro in the central nervous system and segmentation of the Drosophila embryo », Developmental Biology, , p. 209–223 (lire en ligne)
- (en) Nipam H. Patel, Brenda Schafer, Corey S. Goodman, l and Robert Holmgren, « The role of segment polarity genes during Drosophila neurogenesis », Gene and Development by Cold Spring Harbor Laboratory Press, , p. 3:890-904 (ISSN 0890-9369)
- (en) « Arthropod-like Expression Patterns of engrailed and wingless in the Annelid Platynereis dumerilii Suggest a Role in Segment Formation », Current Biology, , Vol. 13, 1876–1881 (lire en ligne, consulté le )
- (en) Nicolas Dray, « Hedgehog Signaling RegulatesSegment Formation in the Annelid Platynereis », Science, , VOL 329