Système Lewis
Le système Lewis, noté LE, no 007 dans la nomenclature de l'ISBT, est un système de groupe sanguin et tissulaire humain.Dans le plasma, les antigènes Lewis sont portés par des glycosphingolipides issus des tissus périphériques. Ils sont adsorbés sur les érythrocytes et deviennent ainsi des déterminants de groupe sanguin. Ils sont portés par les chaînes oligosaccharides de type 1, Galβ1-3GlcNAcβ1-R, des glycoprotéines et des glycosphingolipides dans les épithéliums intestinaux et pulmonaires et dans les glandes exocrines. Ceci est le système Lewis a/b.
Il existe un second système, le système Lewis x/y, qui est basé sur les oligosaccharides de type 2, Galβ1-4GlcNAcβ1-R.
Chaque système dépend de l'action de deux fucosyltransférases, FUT2 et FUT3 pour le premier, FUT1 et FUT3 pour le second.
Ces antigènes sont formés sur une chaîne
de type 1.
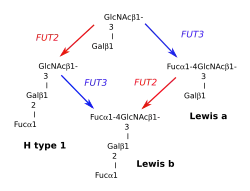
Système Lewis a/b
Génétique
Un premier gène Le, FUT3 pour fucosyltransférase-3, (référence OMIM (en) 111100 ),situé en 19p13.3, produit une α-3/4-L-fucosyltransférase qui fixe un fucose (liaison α1-4) sur la N-acétylglucosamine d'une chaîne Galβ1-3GlcNacβ1-3Galβ1-R de type 1 formant ainsi la substance Lewis a, Lea. L'allèle du gène Le, le, est silencieux. La substance Lea est présente dans les divers liquides biologiques, salive, lait, sperme, urine, intestin, liquide amniotique…
Un second gène, Sécréteur, SE, FUT2, (référence OMIM (en) 182100 ), situé en 19q13.33, produit une α1-2-L-fucosyltransférase qui transforme une chaîne Galβ1-3GlcNacβ1-R en déterminant H de type 1 par adjonction d'un fucose (liaison α1-2) sur le galactose. Le gène SE est exprimé dans les cellules issues de l'endoderme. Son allèle inactif se (mutation G428A), présent en double dose (se/se) chez les sujets non-sécréteurs, ne permet pas cette transformation. Le gène Se est dominant, fréquence génique en Europe 0,52, son allèle se — fréquence 0,48 — est récessif.
Lorsque les deux enzymes sont présentes, FUT2 peut compléter le déterminant Lewis a et conduire au déterminant Lewis b. De même, FUT3 peut compléter H type 1 et donner le déterminant Lewis b.
FUT2 permet la sécrétion des substances A, B ou H (selon le groupe ABO du sujet) dans la salive et autres fluides biologiques, sperme, sécrétions vaginales, gastriques, intestinales, bronchiques, lait, sueur, etc. des sujets sécréteurs (Se/Se ou Se/se). Lorsque le sujet est de plus Le/Le ou Le/le, il peut exprimer ALeb si son groupe sanguin est A ou BLeb si son groupe est B. Ces substances ont été mises en évidence par des techniques d'inhibition d'agglutination.
Phénotypes
Nous observons donc les phénotypes Lewis suivants, selon leurs génotypes :
Le(a-, b-) si le sujet est le/le, qu'il ait le gène Se ou non. Fréquence chez les caucasiens 10 %.
Le(a+, b-) si le sujet est Le/le ou Le/Le, et se/se. Fréquence chez les caucasiens 20 %.
Le(a-, b+) si le sujet est Le/le ou Le/Le, et Se/se ou Se/Se. Fréquence chez les caucasiens 70 %.
Ce dernier phénotype s'explique par le fait que la fucosyltransférase FUT2 (SE) est beaucoup plus active que la fucosyltransférase FUT3. Tous les antigénes Lea produits sont transformés en Leb. Il existe un phénotype Le(a+, b+), dû à un gène Se muté( Ile129Phe), dénommé sej (pour se-Japon), Sew ou Sew385. Ce gène est présent en Asie du Sud-Est (27 % chez les Chinois de Hong Kong), dans la région Pacifique et chez les aborigènes australiens. Il produit une fucosyltransférase de faible activité, qui ne sature par tous les sites Lea.
Leb est le récepteur d'Helicobacter pylori, bactérie responsable d'ulcère, au niveau de l'épithélium gastrique.
Anticorps anti-Lewis
Les anticorps anti-Lea et anti-Leb sont, en règle générale, des anticorps naturels, actifs en technique enzymatique (papaïne) lors de le recherche d'anticorps irrégulier. Ces anticorps n'ont guère, voire pas, d'intérêt transfusionnel.
Il existe divers anticorps anti-Lewis. L'anti-Lea (LE1), l'anti-Leb (LE2), l'anti-Leab, encore appelé anti-Lex (LE3).
L'anti-Lea qui permet, par phénotypage érythrocytaire, de dépister les sujets Le(a+,b-) sans problème.
Il existe deux sortes d'anti-Leb, l'anti-LebL qui dépiste tous les sujets Leb, et l'anti LebH (LE4) qui dépiste préférentiellement les sujets Leb de groupe O ou A2 dont la substance H n'a pas été totalement transformée.
Deux autres anticorps beaucoup plus rares, l'anti-ALeb (anti-LE5) reconnait l'antigène composé A1-Leb, l'anti BLeb (anti-LE6) reconnait l'antigène composé BLeb.
L'anti-Lex (LE3) qui dépiste tous les sujets Le, qu'ils soient Lea ou Leb.
Les anti-LebL et anti-Lex n'étant plus commercialisés, certaines personnes de groupe A1 peuvent parfois être phénotypées Le(a-, b-) avec les anti-Lea et antiLebH dont nous disposons alors qu'en réalité elles sont Le(a-, b+). Ceci a peu d'importance, le système Lewis ne présentant guère d'intérêt transfusionnel.
Il a été décrit des anticorps plasmatiques anti-Lec et anti-Led. Les premiers reconnaissent le disaccharide Galb1-3GlcNAcb1-, précurseurs des chaînes de type 1. Les seconds sont des anti-H, précurseurs immédiats des antigènes A ou B de type 2.
Système Lewis et grossesse
30% des femmes Le(a-, b+) ou Le(a+, b-) perdent pendant leur grossesse l'antigène Lewis qu'elles possèdent. Elles apparaissent donc comme Le(a-, b-) et développent un anticorps naturel anti-Lewis, anti-Lea, anti-Leb et/ou anti-Lex. Un mois à six semaines au plus après l'accouchement, cet anticorps a disparu et ces femmes ont retrouvé leur phénotype Lewis normal. Cette perte d'antigène est sans conséquence pour l'érythrocyte, car la substance Lewis est une substance (glycosphingolipide) qui n'appartient pas à la membrane de l'érythrocyte, mais est une substance soluble (que l'on trouve dans le plasma, la salive, les larmes, le lait, le sperme...) adsorbée passivement sur l'érythrocyte.
La substance Lewis n'est pas détectée sur l'érythrocyte du fœtus ni du nouveau-né qui est donc Le(a-, b-) à la naissance. Il apparaît Le(a+, b-) à l'âge d'un mois environ, puis Le(a+, b+) avant de devenir Le(a-, b+) vers l'âge de deux ans si tel doit être son phénotype définitif, lorsqu'il est génétiquement Le (gène Le) et Sécréteur (gène Se, référence OMIM (en) 182100 ), du moins chez les caucasiens. Ceci explique, entre autres raisons, que les anti-Lewis développés chez la mère n'ont aucune conséquence pour le fœtus.
Système Lewis et infection virale
Il a été montré par les équipes de Jacques Le Pendu à Nantes (France) et de Ralph Baric à Chapel Hill (NC, USA) que les déterminants Lewis b portés par les glycoprotéines à la surface des cellules intestinales des sujets sécréteurs étaient des récepteurs des norovirus, ces virus très contagieux qui provoquent des gastro-entérites, principalement dans les maisons de retraites et les navires de croisière. Il s'avère donc que les sujets non-sécréteurs, n'exprimant pas cet antigène à la surface des leurs cellules intestinales, bénéficient d'une protection contre l'infection par ce type de virus[1].
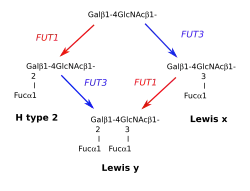
Système Lewis x/y
Le second système Lewis est construit sur les oligosaccharides terminés par une chaîne de type 2. On rencontre les chaînes de type 2 dans les cellules issues du mesoderme ou de l'ectoderme. La biosynthèse des antigènes Lex et Ley est semblable à celle des antigènes Lea et Leb, nécessitant la présence de deux fucosyltransférases.
La première, désignée FUT1, est celle qui permet la synthèse du déterminant H des groupes sanguins ABO. Elle est codée par le gène H ou FUT1 (référence OMIM (en) 211100 ), étroitement lié à Se, ou FUT2 sur le chromosome 19. La seconde est FUT3, la même enzyme que celle qui produit les déterminants Lea et Leb. Elle a la propriété de pouvoir transférer le résidu L-fucose sur le carbone 3 ou le carbone 4 de la N-acétyl-glucosamine, selon que l'un ou l'autre est libre d'occupation.
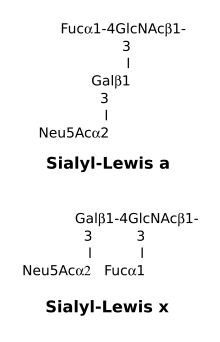
Antigènes Lewis sialylés
Les déterminants Lea et Lex peuvent être sialylés au lieu d'être fucosylés. Dans ce cas l'acide sialique ou acide N-acétylneuraminique est fixé par une sialyltransférase au carbone 3 du D-galactose.
Notes et références
- (en) Lindesmith L, Moe C, Marionneau S, Ruvoen N, Jiang X, Lindblad L, Stewart P, LePendu J and Baric R, « Human susceptibility and resistance to Norwalk virus infection », Nat Med, vol. 9, no 5, , p. 548-553
Voir aussi
Bibliographie
- (en) Human blood groups, Geoff Daniels, Blackwell Science Ltd, 3e édition, 2013.
- (en) The blood group antigen, Marion E. Reid, Christine Lomas-Francis et Martin L. Olsson, Facts Book, Elsevier Academic Press, 3e édition, 2012.
- Les analyses immunohématologiques et leurs applications cliniques, J.Chiaroni, F. Roubinet, P. Bailly, L.Mannessier, F. Noizat-Pirenne, Edit. : John Libbey Eurotext. 2011
- Les groupes sanguins érythrocytaires, coordonné par P. Bailly, J. Chiaroni, F. Roubinet. Edit. : John Libbey Eurotext, Paris, 2015.
Liens externes
- (en) International Society for Blood Transfusion (ISBT)