Satyre élégant
Mutinus elegans
.jpg.webp)
| Règne | Fungi |
|---|---|
| Division | Basidiomycota |
| Classe | Agaricomycetes |
| Sous-classe | Phallomycetidae |
| Ordre | Phallales |
| Famille | Phallaceae |
| Genre | Mutinus |
- Caromyxa elegans Mont. (1856)
- Caryomyxa elegans Mont. (1856)
- Corynites elegans Mont. (1856)
Mutinus elegans, en français Satyre élégant ou Phalle élégant, est un champignon appartenant à la famille des Phallaceae.
Taxinomie et nomenclature
Sous le basionyme de Corynites elegans, il est décrit pour la première fois par le mycologue explorateur français Camille Montagne en 1850 (herbier inédit), publié en 1856 dans son ouvrage Sylloge Cryptogamarum[1].
L'espèce est ensuite recombinée dans le genre Mutinus par Eduard Fischer dans l'ouvrage de Saccardo, Sylloge Fungorum en 1888[2].
Noms savants et vernaculaires

Les noms de genre Mutinus et Ityphallus sont tirés du nom latin de la divinité romaine Mutinus, correspondant à Priape dans la mythologie Grecque, caractérisée par un phallus (pénis symbolique) en érection constante[3]. On appelle Ithyphalle une amulette phallique, portée autour du cou chez les Romains, à fonction apotropaïque, contre le mauvais œil. De même que les phallus, parfois ailés ou munis de clochettes (tintinnabulum), que les Romains suspendaient à l'entrée des maisons, dont on peut voir des exemples au Musée archéologique national de Naples.
Mutinus elegans, sur un plan littéraire, « rebelle, mutin », en raison de la force suggestive de son érection, qui attente à la pudeur[4], et elegans, sans doute en raison de ses belles couleurs .
- Anglais : Elegant stinkhorn, Dog stinkhorn, Devil's dipstick.
- Japonais : タヌキノベニエフデ, 狸紅絵筆 (prononcé « Tanuki-no-béni-éfudé » signifie « Pinceau rouge de blaireau ») [5] - [6].
- C'est également l'étymologie retenue par Imazeki pour le mot také (= « champignons en général », dans les mots composés): Také 菌, 茸, 蕈. Ce mot serait, selon le Daigenkaï, l'abbrévation de takéri, ancien terme pour désigner l'organe mâle, association que l'on retrouve dans de nombreuses civilisations. Takéri est dérivé du verbe takéru, deux mots tombés en désuétude. Takéru a survécu dans les départements de Hiroshima, Shimané, Yamaguchi, Gifu, Tokushima, Ôtsu et Fukuoka avec le sens de « crier » ou « élever la voix, parler à haute voix »; à Mié, Wakayama, Ôsaka, Tokushima et Kôchi sous le sens de « croître, grandir, pousser »; à Niigata et Mié au sens de « gronder, sermonner », et à Miyagi signifiant « copuler, être en rut ». Takéru semble avoir été une variante du verbe taku, signifiant « croître, pousser haut, atteindre la croissance optimale », mais aussi « devenir furieux, rugir, exceller », et cætera. Dans certains usages contemporains, také signifie aussi « haut ». Tous ces sens ont en commun l'idée de « vitalité vigoureuse ». Le mot japonais désignant le bambou est également také, et les philologues nous disent qu'il a la même origine, bien que s'écrivant différemment en caractères sino-japonais 竹. On comprend aisément pourquoi le bambou, à croissance rapide, a été nommé ainsi. Quand les caractères chinois ont été introduits au Japon, le champignon také et le bambou také ont été différenciés en leur attribuant des caractères distincts. Také est un mot plus ancien que kinoko. On trouve le premier dans le Shinsen Jikyô, le plus ancien dictionnaire de Kanjis existant dans son intégralité, publié vers l'an 900 A.D. dans le traité d'herboristerie Honzô Wamyô, ainsi que dans le lexique classifié Wamyô Ruishûshô , publié quelques années plus tard, aucun de ces deux ouvrages ne mentionnant le mot kinoko. Také pour le bambou est attesté dans le Kojiki, qui fut achevé en 712 A.D., et apparaît également dans le Wamyô Ruishûshô. En japonais standard, on trouve také combiné dans les mots composés formant des noms d'espèces de champignons, comme matsu-také (= « champignon du pin ») ou hatsu-také (= « champignon précoce »). D'après le dictionnaire complet des dialectes du Japon, de Misao Tōjō (1951), le terme est encore utilisé seul pour désigner les champignons en général, dans les départements de Nara, Tottori, Hyôgo, Shimané, Ehimé, et Ôïta[6].
Description
.jpg.webp)
.jpg.webp)
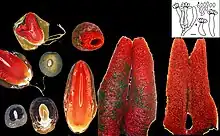
- Champignon d'abord en forme d'œuf, semi-hypogé puis épigé, de la taille d'un œuf de pigeon, subglobuleux puis ovoïde à piriforme, 1,5-4(5) cm de haut x 1-2,7 cm de diamètre, non bosselé mais bientôt allongé par l'érection interne avant l'éclosion; surface lisse, blanche ou partiellement tachée de brunâtre, munis à la base de nombreux rhizomorphes. Péridium constitué de trois couches, les couche externe et interne membraneuses et blanches, et la couche médiane mucilagineuse et hyaline. L'enveloppe externe finit par se rompre pour former une volve à 2-4 lobes, tandis que la couche interne laisse souvent de fines veines blanches, formant des stries ou des bracelets sur le réceptacle[7] - [8].
- Zone fertile non capitée et sans délimitation nette (pas de renflement ni amincissement notable, pas d'alvéoles). Comme chez toutes les Phallales, le réceptacle porte à son sommet (1/4 à 1/3 supérieur) une gléba de couleur vert wagon à brun olivâtre sombre, ointe sur 4-5 cm de la partie fertile, mucilagineuse et d'odeur forte et fétide, parfois à relent douceâtre-nauséeux, mais non cadavérique [7] - [8].
- Réceptacle stipitiforme, 9-19 (22) cm de haut sur 1-2,5 (3) cm de diamètre, de forme très variable, courtement acuminé à la sortie de l’œuf, puis cylindrique, souvent atténué à partir de la mi-hauteur en col de bouteille, toujours appointie et tronquée au sommet, perforée chez l'adulte, le méat du manchon apical variant en diamètre de 0,4-0,8 cm. Structure spongieuse tubulaire et creuse de 2-3 mm d'épaisseur, d'abord dressée comme un i, bientôt courbée, à la fin pliée en deux avant de s’effondrer. Surface entièrement de couleur rouge pastèque caractéristique (rouge carmin orangé, à rouge rose, Methuen 9A4-6, 9A-B7 ou 6A6, 7A7), pâlissant un peu à la base (9C-D8), poreuse dans la partie inférieure qui est composée de cellules polygonales ouvertes de 0,2-0,8 mm, devenant de plus en plus petites et fermées, verruqueuse, vers le haut [7].
- Microscopie : Basidiospores verdâtre pâle dans l'eau, lisses, ellipsoïdes, 4-4,5(5) x 1,8-2 µm, à parois épaisse vers 0,5 µm, non guttulées. Une étude de 1982 a révélé que les spores de Phallaceae, y compris Mutinus elegans, présentent une cicatrice hilaire (0,2–0,3 µm diamètre) visible au microscope électronique, sous forme d'indentation circulaire à un bout de la spore, résultant probablement de la séparation de la spore du stérigmate[9]. Stérigmates de 1,5 µm de long; Basides en massue ou fusiformes, 8-spores, non bouclées, 3-5,5(5) x 20-25(30) µm, se rétrécissant avec l'âge, de sorte que leur largeur est inversement proportionnelle à la taille du primordium étudié, se liquéfiant complètement à maturité pour participer à la formation de la gléba. Cloisons fréquentes, parfois énormes. Hyphes grêles, vers 3 µm, brusquement renflées en raccord, de type manchon de 8 µm. Sphérocystes 20-30 x 25-32 µm. Cystides absentes [7] - [8].
- Écologie : Comme d'autres phallacées, le satyre élégant est à la fois tardif (nov.-déc.), héliophile (recherche la lumière) et thermophile. Il semble affectionner les débris végétaux, surtout les composts d'écorces, et les sols acides, le bord des ruisseaux, les terrains incultes sablonneux ou siliceux. Au Japon, il est apparu pour la première fois en 1999 et quatre années de suite en novembre et décembre à Takatsuki (département d'Osaka) dans la litière des feuilles de bambous, de Quercus, et hors des bois dans les allées herbeuses sous Zelkova et Cinnamomum [7].
- Comestible à l'état d'œuf[7].
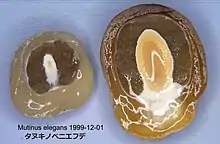
Coupe d'un œuf de 2,5 x 2,5 cm, blanc sordescent, vite sali de gris brunâtre. Se perce naturellement au sommet après deux jours et laisse échapper un, puis deux ballons remplis de mucus translucide. Coupé en deux au cutter, le cuir mince de l'enveloppe externe offre quelque résistance tandis qu'en dessous, une matière visqueuse comme une huître fuit sous la lame, avant de rencontrer une matière plus solide, grenue comme un radis. Un liquide incolore et insipide comme de l'eau s'en échappe[8] - [10].
La coupe (photo ci-contre) montre un cortex rouge rose au centre, plus ou moins cordiforme, presque en V, à parois épaisses de 2-3 mm, enveloppé dans une gélatine translucide, plus colorée opaque qu'un blanc d’œuf, beige à jaune brunâtre pâle (couleur de la gomme arabique, de résine de pin), de 3-6 mm d'épaisseur de part et d'autre de la tige centrale, presque nulle au sommet et à la base[10].
Le réceptacle est creusé en cavité de 2-4 mm sur chaque moitié. La volve est déjà formée par une couche basale blanc pur, isolant le canal de gelée amniotique, comme un nombril de 1,5 mm d'épaisseur, reliée à l'extérieur à deux cordons mycéliens[10].
Le réceptacle de l'adulte présente presque toujours un orifice sommital, foramen de 1-3 mm de diamètre. Le tube est un tissu formé d'alvéoles de 0,5-1 mm (mesurées à la face interne du tube) sur la partie fertile, de plus en plus larges vers la base[10].

Comme chez toutes les espèces de l'ordre des Phallales, le stipe porte à son sommet une gleba mucilagineuse et fétide. Il se différencie du banal satyre du chien (Mutinus caninus) par sa taille plus robuste, appointie au sommet, sa partie fertile colorée de rouge écarlate et peu différenciée du réceptacle (on n'ose pas dire pied, vu l'absence de chapeau ). ce dernier n'ayant pas d'alvéoles sur la face externe, est simplement ponctué de pores [7] - [10].
Confusions fréquentes
Les espèces du genre Mutinus d'Europe et d'Eurasie sont souvent confondues faute de clé d'identification. Les États-Unis et l'Asie de l'Est confondent également souvent M. caninus et M. ravenelii (Fleger et Hooper 1980). Certains mycologues ont même douté de l'existence de M. ravenelii, la considérant parfois comme une simple variété d'un rouge plus intense (Calonge 1990 :46). La littérature européenne de la fin du XXe siècle a souvent manqué d'illustrations valables et de matériel frais à la base de descriptions (Høiland 1977 : 158 fig.1) par exemple, et aux États-Unis, où dans Mushrooms from North America, (Phillips 1991[11]), M. ravenelii est considéré une forme à stipe rose de M. elegans[12].
À condition d'avoir en main des spécimens mûrs:
- M. elegans se distingue par la partie fertile de forme non capitée et par la couleur uniforme. Basides 8-spores.
- M. bambusinus par la partie fertile capitée, acuminée très allongée distincte de la base stérile par sa couleur plus intense (tropicale).
- Mutinus caninus est "capité" par une courte "tête" adhérente, nettement délimitée par un renflement et une dépression en forme d'amande, contrastant aussi par sa couleur rouge orangé sous la gléba[13] - [14]. Odeur faible, de tabac et des basides 6-spores.
- M. ravenelii a une tête courte écarlate carminée très pointue, et le réceptacle d'un rose framboise intense. Odeur urineuse. Basides 8-pores.
Propriétés Antibiotiques
Une étude portant sur 32 champignons basidiomycètes a établi que Mutinus elegans était la seule espèce capable d'activité à la fois antibactérienne et antifongique contre six des micro-organismes testés, nommément les bactéries à effet pathologique chez l'homme : Bacillus cereus, Bacillus subtilis, Staphylococcus aureus, Escherichia coli, Salmonella typhimurium et la levure Candida albicans[15] - [16].
Notes
- page 281 du Sylloge generum specierumque cryptogamarum quas in variis operibus descriptas iconibusque illustratas, nunc ad diagnosim reductas, multasque novas interjectas, ordine systematico disposuit. Paris, Baillière. 1856, XXIV, (498pp.)
- Saccardo, P.A. 1888. Sylloge Fungorum. 7 p. 13
- Arora, David., Mushrooms demystified : a comprehensive guide to the fleshy fungi, Ten Speed Press, (ISBN 0-89815-170-8, 978-0-89815-170-1 et 0-89815-169-4, OCLC 13702933, lire en ligne)
- 4 siècle après J.C. Lactantius(Lactance) Mutunus n. : dieu de la virilité représenté par un phallus
- Les personnages légendaires ou mythologiques que sont le Tanuki (blaireau) et le kitsuné (renard) sont traditionnellement utilisés en sciences naturelles dans les noms savants des espèces spectaculaires ou inquiétantes, souvent pour traduire une connotation érotique. Le tanuki est réservé aux espèces de grandes tailles.
- 今関六也 Rokuya Imazeki 1973 - Japanese mushroom names (Transactions of the Asiatic Society of Japan, 3rd ser., v. 11, pp. 25-80) Lire en ligne (traduction française)
- Guez, D. & Nagasawa, E. (2000) - Trans. mycol. Soc. Japan 42 : 75-78 (ja) (résumé en anglais) Lire en ligne
- Marchand, A. (1976). - Champignons du Nord et du Midi, tome 4. Aphyllophorales (fin), Hydnaceae, Gasteromycetes, Ascomycetes. 263 pp., pl. 378, Ed. Société mycologique des Pyrénées méditerranéennes, Diffusion Hachette, (ISBN 84-399-4768-2)
- William R. Burk, Stanley L. Flegler et Wilford M. Hess, « Ultrastructural Studies of Clathraceae and Phallaceae (Gasteromycetes) Spores », Mycologia, vol. 74, no 1, , p. 166 (DOI 10.2307/3792646, lire en ligne, consulté le )
- Phalle élégant, Mutin élégant (タヌキノベニエフデ) Lire en ligne
- Phillips, R., (1991). - Mushrooms of North America, Little, Brown & Co., Boston, (319 pp.), pl. p. 289 (gauche);
- Szczepka, M.Z. - 1995 Clé d'identification du genre Mutinus Fr. en Europe Doc. mycol. XXV fasc. 98-100 p.453-458
- Jean Mornand (1984) - Gastéromycètes de France, Doc. Mycol. tome 14, fasc. 53 p. 41-50, p. 43
- Marchand André (1976) : Champignons du nord et du midi, Tome 4, Aphyllophorales, hydnaceae, gastéromycètes, (ISBN 84-399-4768-2) ; no. 377
- Bianco Coletto MA, Lelli P (1998–99). "Antibiotic activity in Basidiomycetes. XII. Antibacterial and antifungal activity of 32 new strains". Allionia (Turin). 36: 89–92.
- Bianco Coletto MAB (2005–06). "Antibiotic activity of some new recently isolated strains". Allionia (Turin). 40: 33–7.
Bibliographie
- Lloyd, C.G. (1906). - Synopsis Phalloideae Mycological notes no.24., Bull. Trim. Soc. Mycol. France 23 : 244-246;
- Guéguen, F., 1908 (1907). - Bibliographie analytique;
- Marchand, A. (1976). - Champignons du Nord et du Midi, tome 4. Aphyllophorales (fin), Hydnaceae, Gasteromycetes, Ascomycetes. 263 pp., pl. 378, Ed. Société mycologique des Pyrénées méditerranéennes, Diffusion Hachette, (ISBN 84-399-4768-2) ;
- Cetto B., (1978). - I funghi dal Vero, Saturnia, Trento, Vol. 2 (3e édition), pl. 798;
- Lincoff G.H., (1981). - National Audubon Society Field Guide to North American Mushrooms. A. Knopf, New York, (926 pp.), : 692;
- Benzoni, C. (1981). - Bull. Suisse Mycol. 59(9) :146 ;
- Arora, D. (1986). - Mushrooms Demystified, (2e édition), Ten Speed Press, Berkeley, (959 pp.) : 195-196;
- Phillips, R., (1991). - Mushrooms of North America, Little, Brown & Co., Boston, (319 pp.), pl. p. 289 (gauche);
- Bulletin de la Fédération Mycologique Dauphiné-Savoie 71 (couverture);
- Rivista de Micologia (1992) (1) : 41;
- Montégut, J. (1992). - Encyclopédie analytique des champignons. Vol. 1, Ed. SECN, Orgeval. pl. 186 ;
- Szczepka, M.Z. - 1995 Clé d'identification du genre Mutinus Fr. en Europe, Doc. mycol. XXV fasc. 98-100 p.453-458
- Guez, D. & Nagasawa, E. (2000) - Trans. mycol. Soc. Japan 42 : 75-78 (ja) (résumé en anglais) Lire en ligne