Intégration de chemin
L'intégration de chemin, ou intégration de trajet, est définie comme la réactualisation d'une représentation interne de la position et de la direction d'un sujet dans l'environnement en se basant sur des informations du corps en mouvement[1].
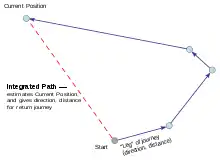
De nombreux animaux retournant à leur résidence entre deux périodes de recherche de nourriture ont recours à l'intégration de chemin, notamment des mammifères (dont des rongeurs) et des arthropodes (hyménoptères sociaux, araignées, crabes violonistes, mantes de mer)[2].
Historique
En 1873, Charles Darwin émet pour la première fois l'hypothèse d'un système de navigation par inertie chez les animaux[3]. Puis en 1913, les travaux de Trowbridge introduisent pour la première fois la notion de carte cognitive spatiale[4]. À partir du milieu du XXe siècle, Tolman et d'autres chercheurs ont montré que les animaux pouvaient retourner directement à un point de départ, comme pour retrouver leur nid, et ce même dans l'obscurité[5]. Cela suggère que les animaux sont capables d'utiliser des repères spécifiques pour calculer la distance et leur direction afin d'estimer leur position, et donc de savoir comment revenir à leur position initiale. Ce processus a été nommé intégration de chemin car il permet de saisir le concept d'intégration continue des repères liés au mouvement tout au long d'un parcours.
La notion d'intégration de chemin chez les mammifères répond à trois découvertes majeures. La première, au début des années 1970, indique que l'activité neuronale hippocampique du rat est corrélée à la position de la tête de l'animal dans l’espace[6]. Cela a conduit à la découverte de neurones spécifiques situés dans l'hippocampe : les cellules de lieu qui répondent à la position de l'animal[7].
La seconde découverte, au début des années 1990, est celle des « cellules de direction de la tête », qui répondent à l'orientation de la tête de l'animal[8]. Ce type de neurones se situe au niveau du thalamus ainsi que dans le subiculum[9]. Les cellules de direction de la tête peuvent être qualifiées de « boussoles neurales »[10]. En effet, l'activation de ces neurones se fait par émission de potentiels d'action qui sont définis par rapport à une représentation interne de l’espace qui se construit au fur et à mesure des déplacements de l'animal dans un nouvel environnement[11].
La troisième découverte, au début du XXIe siècle, est celle de neurones spécifiques au sein du cortex entorhinal dorso-médial (dMEC) : les cellules de grille[12]. L'émission de potentiel d'actions associés aux cellules de grille révèlent un motif particulier: un maillage régulier hexagonal apparaît lorsqu'un sujet se déplace dans l'environnement.
Néanmoins, il reste encore à savoir si les cellules de lieu et de grille calculent réellement un signal d'intégration de chemin, mais il existe des modèles de calcul qui suggèrent que cela est plausible. Il est certain que les lésions cérébrales dans ces régions semblent nuire à la capacité des animaux à s'intégrer.
Mécanisme
Le mécanisme d'intégration de chemin intervient dans les opérations de mise à jour spatiale de la position et de l'orientation de l'animal par rapport à son point de départ. Les déplacements de l'animal sont basés sur les mouvements de son corps :
- soit translationnels, qui correspondent à un mouvement linéaire du corps et de la tête,
- soit rotationnels en fonction des modifications de direction du corps et de la tête.
L'intégration de chemin est donc un processus dynamique de mémorisation spatiale de l'environnement qui permet de corriger les erreurs accumulées lors du trajet tout en connaissant la distance et la direction de son point de départ. Pour revenir à son point de départ, l'animal n'a pas nécessairement besoin de mémoriser l'ensemble du trajet parcouru, ni même de retenir les points d’arrêts effectués. Ainsi, les seules données enregistrées par l'animal sont celle de son emplacement courant et celle de son point de départ, qui au cours de son trajet sont continuellement réactualisées[13].
La plupart des modèles de navigation dans un environnement nécessitent d'intégrer des informations précises : sélectionner des chemins, de planifier et d'exécuter un trajet d'une position initiale à une position finale. Pour cela, les humains utilisent deux stratégies distinctes pour la navigation : d'une part la navigation « allocentrique » et d'autre part « égocentrique »[14]. La stratégie qualifiée d'égocentrique englobe des informations du monde extérieur. Celle-ci repose sur l'utilisation de représentations de type « stimulus-réponse » qui concernent le changement de direction du corps associé à un stimulus particulier. Ce dernier est lié au fonctionnement du noyau caudé. Au sujet de la stratégie de navigation allocentrique, celle-ci s'appuie sur les informations internes de notre système. Elle reposerait principalement sur l'élaboration d'une carte cognitive associée au fonctionnement de l'hippocampe et plus particulièrement de la partie du cortex entorhinal située en dessous de l'hippocampe[15]. Parmi les informations internes, une partie provient du système vestibulaire à la base de notre sens de l'équilibre et de notre sensation du mouvement. Ces informations qualifiées de vestibulaires génèrent au niveau de l'hypothalamus des informations directionnelles qui sont codées par des neurones spéciaux appelés « cellules de direction de la tête ». D'autres sources d'informations internes sont la proprioception, via les efférences motrices qui correspondent à l'information du système moteur indiquant au reste du cerveau quels mouvements ont été commandés et exécutés, et le flux optique[16]. Réunies, ces sources d'informations peuvent indiquer à l'animal dans quelle direction il se déplace, à quelle vitesse et sur quelle durée. De plus, des informations telles que la sensibilité au champ magnétique terrestre pour les animaux souterrains (par exemple : les rats taupes) peuvent donner une intégration de trajectoire[17].
Voir aussi
Références
- H. Mittelstaedt et M.-L. Mittelstaedt, « Homing by Path Integration », dans Avian Navigation, Springer Berlin Heidelberg, (ISBN 978-3-642-68618-4, DOI 10.1007/978-3-642-68616-0_29, lire en ligne), p. 290–297.
- (en) Rickesh N. Patel et Thomas W. Cronin, « Mantis Shrimp Navigate Home Using Celestial and Idiothetic Path Integration », Current Biology, vol. 30, no 11, , p. 1981-1987.e3 (DOI 10.1016/j.cub.2020.03.023
 ).
). - (en) Charles Darwin, « Origin of Certain Instincts », Nature, vol. 7, no 179, , p. 417–418 (ISSN 1476-4687, DOI 10.1038/007417a0, lire en ligne, consulté le ).
- (en) C. C. Trowbridge, « ON FUNDAMENTAL METHODS OF ORIENTATION AND "IMAGINARY MAPS" », Science, vol. 38, no 990, , p. 888–897 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.38.990.888, lire en ligne, consulté le ).
- Edward C. Tolman, « Cognitive maps in rats and men. », Psychological Review, vol. 55, no 4, , p. 189–208 (ISSN 1939-1471 et 0033-295X, DOI 10.1037/h0061626, lire en ligne, consulté le ).
- (en) J. O'Keefe et J. Dostrovsky, « The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat », Brain Research, vol. 34, no 1, , p. 171–175 (DOI 10.1016/0006-8993(71)90358-1, lire en ligne, consulté le ).
- John O'Keefe et Lynn Nadel, « The cognitive map as a hippocampus », Behavioral and Brain Sciences, vol. 2, no 4, , p. 520–533 (ISSN 0140-525X et 1469-1825, DOI 10.1017/s0140525x00064256, lire en ligne, consulté le ).
- (en) Js Taube, Ru Muller et Jb Ranck, « Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis », The Journal of Neuroscience, vol. 10, no 2, , p. 420–435 (ISSN 0270-6474 et 1529-2401, PMID 2303851, PMCID PMC6570151, DOI 10.1523/JNEUROSCI.10-02-00420.1990, lire en ligne, consulté le ).
- (en) J. S. Taube, « Persistent Neural Activity in Head Direction Cells », Cerebral Cortex, vol. 13, no 11, , p. 1162–1172 (ISSN 1460-2199, DOI 10.1093/cercor/bhg102, lire en ligne, consulté le ).
- (en) Robert U Muller, James B Ranck et Jeffrey S Taube, « Head direction cells: properties and functional significance », Current Opinion in Neurobiology, vol. 6, no 2, , p. 196–206 (DOI 10.1016/S0959-4388(96)80073-0, lire en ligne, consulté le ).
- (en) Jean Simonnet, Mérie Nassar, Federico Stella et Ivan Cohen, « Activity dependent feedback inhibition may maintain head direction signals in mouse presubiculum », Nature Communications, vol. 8, no 1, , p. 16032 (ISSN 2041-1723, PMID 28726769, PMCID PMC5524997, DOI 10.1038/ncomms16032, lire en ligne, consulté le ).
- (en) Torkel Hafting, Marianne Fyhn, Sturla Molden et May-Britt Moser, « Microstructure of a spatial map in the entorhinal cortex », Nature, vol. 436, no 7052, , p. 801–806 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/nature03721, lire en ligne, consulté le ).
- Séguinot, Valérie, L'intégration du trajet: approches théorique et expérimentale chez le hamster et le chien. (OCLC 631960172, lire en ligne).
- Mathieu Taillade, Bernard N’Kaoua, Prashan Arvind Pala et Hélène Sauzéon, « Cognition spatiale et vieillissement : les nouveaux éclairages offerts par les études utilisant la réalité virtuelle », Revue de neuropsychologie, vol. 6, no 1, , p. 36 (ISSN 2101-6739 et 2102-6025, DOI 10.3917/rne.061.0036, lire en ligne, consulté le ).
- (en) Torkel Hafting, Marianne Fyhn, Sturla Molden et May-Britt Moser, « Microstructure of a spatial map in the entorhinal cortex », Nature, vol. 436, no 7052, , p. 801–806 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/nature03721, lire en ligne, consulté le ).
- (en) Bruce L. McNaughton, Francesco P. Battaglia, Ole Jensen et Edvard I Moser, « Path integration and the neural basis of the 'cognitive map' », Nature Reviews Neuroscience, vol. 7, no 8, , p. 663–678 (ISSN 1471-003X et 1471-0048, DOI 10.1038/nrn1932, lire en ligne, consulté le ).
- T. Kimchi, A. S. Etienne et J. Terkel, « A subterranean mammal uses the magnetic compass for path integration », Proceedings of the National Academy of Sciences, vol. 101, no 4, , p. 1105–1109 (ISSN 0027-8424 et 1091-6490, DOI 10.1073/pnas.0307560100, lire en ligne, consulté le ).
Bibliographie
- (en) Bruce L. McNaughton, Francesco P. Battaglia, Ole Jensen et Edvard I Moser, « Path integration and the neural basis of the 'cognitive map' », Nature Reviews Neuroscience, vol. 7, no 8, , p. 663–678 (ISSN 1471-003X et 1471-0048, DOI 10.1038/nrn1932).
- (en) Torkel Hafting, Marianne Fyhn, Sturla Molden et May-Britt Moser, « Microstructure of a spatial map in the entorhinal cortex », Nature, vol. 436, no 7052, , p. 801–806 (ISSN 0028-0836 et 1476-4687, DOI 10.1038/nature03721).
- Séguinot, Valérie, L'intégration du trajet: approches théoriques et expérimentales chez le hamster et le chien. (OCLC 631960172 ).