Diauxie
La diauxie est un mot d'origine grec inventé par Jacques Monod pour désigner une croissance à deux phases[1]. Le mot est utilisé en anglais (diauxie) en biologie cellulaire pour décrire les phases de croissance d'un microorganisme en culture discontinue (batch) puisqu'il métabolise un mélange de deux sucres. Plutôt que de métaboliser simultanément les deux sucres disponibles, les cellules microbiennes les consomment généralement selon un schéma séquentiel, ce qui entraîne deux phases de croissance distinctes.
Phases de croissance
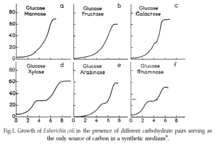
Jacques Monod a découvert la croissance diauxique en 1941 lors de ses expériences avec Escherichia coli et Bacillus subtilis. Tout en cultivant ces bactéries sur différentes combinaisons de sucres au cours de sa thèse de doctorat, Monod a observé que souvent, deux phases de croissance distinctes sont clairement visibles dans la culture en batch comme le montre l'image ci-contre.
Au cours de la première phase, les cellules métabolisent préférentiellement le sucre sur lequel elles peuvent croître plus rapidement (souvent du glucose mais pas toujours). Une fois le premier sucre épuisé, les cellules passent au second. Au moment du "décalage diauxique", il existe souvent une période de latence pendant laquelle les cellules produisent les enzymes nécessaires à la métabolisation du second sucre.
Monod a par la suite mis de côté ses travaux sur la croissance diauxique et s'est concentré sur le modèle d'expression génique de l'opéron lac, qui l'a conduit à un prix Nobel[2].
La diauxie se produit parce que les organismes utilisent des opérons ou de multiples ensembles de gènes pour contrôler différemment l'expression des enzymes nécessaires pour métaboliser les différents nutriments ou sucres qu'ils rencontrent. Si un organisme utilise son énergie et d’autres ressources (acides aminés par exemple) pour synthétiser les enzymes nécessaires à la métabolisation d’un sucre qui ne peut que soutenir un taux de croissance plus lent et n'utilise pas la totalité ou la quasi-totalité de ses ressources disponibles pour synthétiser les enzymes qui métabolisent un sucre différent permettant un taux de croissance plus rapide, un tel organisme aura un désavantage reproductif par rapport à ceux qui choisiront d'utiliser le sucre permettant une croissance plus rapide. Au fil de l'évolution, les organismes ont développé la capacité de réguler leurs mécanismes de contrôle génétique de manière à n'exprimer que ces gènes permettant le taux de croissance le plus rapide. Par exemple, lorsqu'elle est cultivée en présence de glucose et de maltose, la bactérie Lactococcus lactis produit des enzymes permettant de métaboliser le glucose en premier, modifiant ainsi l'expression de son gène pour utiliser le maltose uniquement après épuisement de la quantité de glucose disponible.
Dans le cas de la levure Saccharomyces cerevisiae utilisée en boulangerie ou brasserie qui croît en présence de glucose avec beaucoup d'aération, le modèle de croissance diauxique est généralement observé dans la culture batch. Au cours de la première phase de croissance, lorsqu'il y a de grandes quantités de glucose et d'oxygène disponibles, les cellules de levure préfèrent la fermentation du glucose à la respiration aérobie, bien que cette dernière soit la voie la plus efficace pour croître en présence de glucose. Contrairement à l'effet Pasteur plus communément invoqué, ce phénomène de préférence pour la fermentation favorisant une croissance plus rapide est plus proche de l'effet Warburg observé dans les tumeurs à croissance rapide. Les mécanismes de régulation génétique intracellulaires ont évolué pour imposer ce choix, car la fermentation fournit aux cellules de levure un taux de croissance plus rapide que la respiration aérobie du glucose. Une fois le glucose épuisé, l'éthanol produit par fermentation est oxydé en une seconde phase de croissance sensiblement plus lente, si de l'oxygène est disponible.
Mécanisme proposé
Dans les années 1940, Monod a émis l'hypothèse qu'une seule enzyme pourrait s'adapter pour métaboliser différents sucres. Il a fallu 15 ans de travaux supplémentaires pour montrer que cela était incorrect. Au cours de ses travaux sur l'opéron lac chez E. coli, Joshua Lederberg a isolé la β-galactosidase et l'a trouvée en plus grande quantité dans les colonies cultivées sur du lactose par rapport aux autres sucres[3]. Melvin Cohn, collègue de Monod à l'Institut Pasteur, a alors découvert que les β-galactosides induisaient une activité enzymatique[4]. La notion d'adaptation enzymatique a donc été remplacée par la notion d'induction enzymatique, dans laquelle une molécule induit l'expression d'un gène ou d'un opéron, souvent en se liant à une protéine répressive et en l'empêchant de se lier à l'opérateur[5].
Dans le cas de la diauxie bactérienne où l'on passe du métabolisme du glucose à celui du lactose, un mécanisme proposé a suggéré que le glucose inhibe d'abord la capacité de l'enzyme adénylate cyclase à synthétiser l'AMP cyclique (AMPc). L'AMPc, quant à elle, est nécessaire pour que la protéine activatrice de catabolite (CAP) se lie à l'ADN et active la transcription de l'opéron lac, qui comprend les gènes nécessaires au métabolisme du lactose. La présence d'allolactose, un produit métabolique du lactose, est détectée par l'activité du répresseur lac, qui inhibe la transcription de l'opéron lac jusqu'à ce que le lactose soit présent. Ainsi, si le glucose est présent, les taux d'AMPc restent bas, de sorte que la CAP est incapable d'activer la transcription de l'opéron lac, indépendamment de la présence ou de l'absence de lactose. Lorsque les réserves de glucose sont épuisées, les taux d'AMPc augmentent, ce qui permet à la CAP d'activer les gènes nécessaires au métabolisme d'autres sources de nourriture, y compris le lactose, le cas échéant[6].
Des recherches plus récentes suggèrent toutefois que le modèle avec l'AMPc n'est pas correct dans ce cas puisque les niveaux d'AMPc restent identiques dans les conditions de croissance du glucose et du lactose, et un modèle différent a été proposé et suggère que la diauxie lactose-glucose chez E. coli pourrait être provoquée principalement par exclusion de l'inducteur[7]. Dans ce modèle, le transport du glucose via le EIIA Glc arrête la lactose perméase lorsque le glucose est transporté dans la cellule. Le lactose n'est donc pas transporté puis utilisé dans la cellule. Bien que le mécanisme AMPc/CAP puisse ne pas jouer de rôle dans la diauxie glucose/lactose, on pense qu'il agit comme mécanisme dans d’autres cas de diauxie.
Voir aussi
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Diauxie » (voir la liste des auteurs).
Références
- Jacques Monod, « Diauxie et respiration au cours de la croissance des cultures de B. coli », dans Selected Papers in Molecular Biology by Jacques Monod, Academic Press, (ISBN 978-0-12-460482-7, lire en ligne), p. 12–13
- C Burstein, M Cohn, A Kepes et J Monod, « Rôle du lactose et de ses produits métaboliques dans l'induction de l'opéron lactose chez Escherichia coli », Biochimica et Biophysica Acta (BBA) - Nucleic Acids and Protein Synthesis, vol. 95, no 4, , p. 634–639 (ISSN 0005-2787, DOI 10.1016/0005-2787(65)90517-4, lire en ligne, consulté le )
- Joshua Lederberg, « THE BETA-d-GALACTOSIDASE OF ESCHERICHIA COLI, STRAIN K-121 », Journal of Bacteriology, vol. 60, no 4, , p. 381–392 (ISSN 0021-9193, PMID 14784466, lire en ligne, consulté le )
- Melvin Cohn, « CONTRIBUTIONS OF STUDIES ON THE β-GALACTOSIDASE OF ESCHERICHIA COLI TO OUR UNDERSTANDING OF ENZYME SYNTHESIS1 », Bacteriological Reviews, vol. 21, no 3, , p. 140–168 (ISSN 0005-3678, PMID 13471456, lire en ligne, consulté le )
- Mulligan, « Induction » [archive du ] (consulté le )
- Brown, « Transient Changes in Gene Activity » (consulté le )
- Stülke J, Hillen W., « Carbon catabolite repression in bacteria », Current Opinion in Microbiology, vol. 2, no 2, , p. 195–201 (PMID 10322165, DOI 10.1016/S1369-5274(99)80034-4)