ADN ribosomique
L'ADN ribosomique (ADNr) est l'ensemble des séquences d'ADN transcrites en ARN ribosomiques (ARNr). Les ribosomes sont des complexes de protéines et d'ARNr au sein desquels se déroule la traduction des ARN messagers au cours de la biosynthèse des protéines.
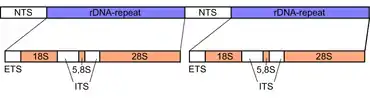
L'ADN ribosomique des eucaryotes est un opéron formé d'une répétition en tandem de domaines NTS, ETS, ARNr 18S, ITS1, ARNr 5,8S, ITS2 et ARNr 28S. Il possède également un autre gène transcrit en ARNr 5S dans le génome de la plupart des eucaryotes[1]. L'ARNr 5S est également présent dans les tandems répétés par exemple chez la drosophile. Dans le noyau, la région du chromosome portant l'ADN ribosomique apparaît sous la forme d'un nucléole, qui forme de grandes boucles d'ADNr. Ces régions d'ADNr sont également appelées organisateur nucléolaire dans la mesure où elles forment la base structurelle du nucléole. Le génome humain compte cinq chromosomes portant de telles régions : les chromosomes acrocentriques 13, 14, 15, 21 et 22.
Conservation de la séquence des unités répétées
Le polymorphisme entre répétitions d'ADN ribosomique reste très limité, ce qui indique une évolution concertée (en) entre les tandems d'ADNr[1]. Le mécanisme de cette évolution concertée n'est cependant pas parfait, de sorte qu'il existe bien des divergences, qui permettent de réaliser des analyses phylogénétiques entre organismes apparentés[2] - [3].
La comparaison de séquences répétées d'ARNr 5S entre plusieurs drosophiles a révélé que les insertions et les délétions sont fréquentes entre les espèces et voisinent souvent avec des séquences conservées[4]. Ces mutations peuvent se produire par glissement du brin d'ADN nouvellement synthétisé lors de la réplication de l'ADN ou par conversion génique[4].
Analyses phylogénétiques dérivées des polymorphismes observés
Les domaines transcrits de l'ADN ribosomique présentent un faible taux de polymorphisme entre les espèces, ce qui permet des comparaisons entre un petit nombre d'individus en vue d'établir leur parenté phylogénétique.
Les espaceurs internes transcrits (ITS) ont un taux de mutations sensiblement plus élevé sous l'effet d'insertions, de délétions et de mutations ponctuelles, ce qui rend inopérante la comparaison de séquences d'ITS entre espèces très éloignées, comme les humains et les grenouilles par exemple[5]. Il est toutefois possible de mener avec succès des analyses phylogénétiques entre espèces voisines en incluant les ITS.
En revanche, l'analyse des régions transcrites de l'ADN ribosomique permet de réaliser de telles études car la séquence de ces régions évolue bien plus lentement. On peut ainsi analyser valablement les divergences entre un humain et une levure : l'ARNr 5,8S de ces deux organismes est identique à 75 %[6].
De cette façon, l'ensemble de l'ADN ribosomique est susceptible de fournir des informations phylogénétiques sur un large éventail d'espèces[7].
Notes et références
- (en) Guy-Franck Richard, Alix Kerrest et Bernard Dujon, « Comparative Genomics and Molecular Dynamics of DNA Repeats in Eukaryotes », Microbiology and Molecular Biology Reviews, vol. 72, no 4, , p. 686-727 (PMID 19052325, PMCID 2593564, DOI 10.1128/MMBR.00011-08, lire en ligne)
- (en) I. Álvarez et J. F. Wendel, « Ribosomal ITS sequences and plant phylogenetic inference », Molecular Phylogenetics and Evolution, vol. 29, no 3, , p. 417-434 (PMID 14615184, DOI 10.1016/S1055-7903(03)00208-2, lire en ligne)
- (en) Kevin Weitemier, Shannon C.K. Straub, Mark Fishbein et Aaron Liston, « Intragenomic polymorphisms among high-copy loci: a genus-wide study of nuclear ribosomal DNA in Asclepias (Apocynaceae) », PeerJ, , e718 (PMID 25653903, PMCID 4304868, DOI 10.7717/peerj.718, lire en ligne)
- (en) F. Pâques, M. L. Samson, P. Jordan et M. Wegnez, « Structural evolution of the Drosophila 5S ribosomal genes », Journal of Molecular Evolution, vol. 41, no 5, , p. 615-621 (PMID 7490776, DOI 10.1007/BF00175820, lire en ligne)
- (en) Masayuki Sumida, Yoji Kato et Atsushi Kurabayashi, « Sequencing and analysis of the internal transcribed spacers (ITSs) and coding regions in the EcoR I fragment of the ribosomal DNA of the Japanese pond frog Rana nigromaculata », Genes & Genetic Systems, vol. 79, no 2, , p. 105-118 (PMID 15215676, DOI 10.1266/ggs.79.105, lire en ligne)
- (en) Ross N. Nazar, Thomas O. Sitz et Harris Busch, « Sequence homologies in mammalian 5.8S ribosomal RNA », Biochemistry, vol. 15, no 3, , p. 505-508 (PMID 1252408, DOI 10.1021/bi00648a008, lire en ligne)
- (en) David M. Hillis et Michael T. Dixon, « Ribosomal DNA: Molecular Evolution and Phylogenetic Inference », The Quarterly Review of Biology, vol. 66, no 4, , p. 411-453 (PMID 1784710, JSTOR 2831326)