Pronucleus
Un pronoyau (pronuclei au pluriel) est le noyau d'un gamète, soit du spermatozoïde, soit de l'ovule pendant le processus de fécondation. Le spermatozoïde devient un pronucléus après l'entrée du spermatozoïde dans l'ovule, mais avant que le matériel génétique du spermatozoïde et de l'ovule ne fusionne. Contrairement au spermatozoïde, l'ovule possède un pronucléus une fois qu'il devient haploïde, et non à l'arrivée du spermatozoïde. Les spermatozoïdes et les ovules sont haploïdes, ce qui signifie qu'ils portent la moitié du nombre de chromosomes des cellules somatiques, donc chez l'homme, les cellules haploïdes ont 23 chromosomes, tandis que les cellules somatiques ont 46 chromosomes. Les pronucléi mâles et femelles ne fusionnent pas, contrairement à leur matériel génétique. Au lieu de cela, leurs membranes se dissolvent, ne laissant aucune barrière entre les chromosomes mâles et femelles. Leurs chromosomes peuvent alors se combiner et faire partie d'un seul noyau diploïde dans l'embryon résultant, contenant un ensemble complet de chromosomes.
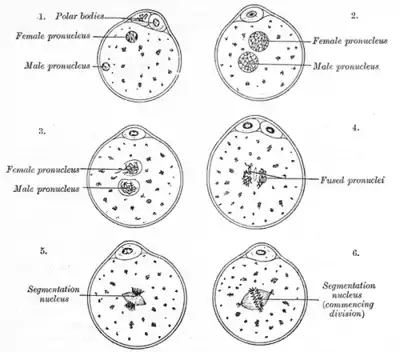
L'apparition de deux pronucléi est le premier signe d'une fécondation réussie telle qu'observée lors d'une fécondation in vitro, et est généralement observée 18 heures après l'insémination ou l'injection intracytoplasmique de spermatozoïdes. L'embryon est alors appelé zygote à deux pronuclei. Les zygotes à deux pronuclei passant par les états à un pronoyau ou trois pronuclei ont tendance à se développer en embryons de moins bonne qualité que ceux qui restent à deux pronuclei tout au long du développement [1] et peuvent être importants dans la sélection des embryons lors de la fécondation in vitro.
Histoire
Le pronoyau a été découvert dans les années 1870 au microscope en utilisant des techniques de coloration combinées à des microscopes avec des niveaux de grossissement améliorés. Le pronoyau a été trouvé à l'origine lors des premières études sur la méiose. Édouard Van Beneden a publié un article en 1875 dans lequel il mentionne pour la première fois le pronucléus en étudiant les œufs de lapins et de chauves-souris. Il a déclaré que les deux pronoyaux se forment ensemble au centre de la cellule pour former le noyau embryonnaire. Van Beneden a également découvert que le spermatozoïde pénètre dans la cellule à travers la membrane pour former le pronucléus mâle. En 1876, Oscar Hertwig a fait une étude sur les œufs d'oursins parce que les œufs d'oursins sont transparents, ce qui a permis un bien meilleur grossissement de l'œuf. Hertwig a confirmé la découverte du pronucléus par Van Beneden et a également découvert que la formation du pronucléus féminin implique la formation de corps polaires[2].
Formation
Le pronucléus femelle est l'ovule femelle une fois qu'il est devenu une cellule haploïde, et le pronucléus mâle se forme lorsque le spermatozoïde pénètre dans l'ovule femelle. Alors que le sperme se développe à l'intérieur des testicules mâles, le spermatozoïde ne devient pas un pronucléus jusqu'à ce qu'il se décondense rapidement à l'intérieur de l'ovule femelle[3]. Lorsque le spermatozoïde atteint l'ovule femelle, le spermatozoïde perd sa membrane extérieure ainsi que sont flagelle. La formation de l'ovule femelle est asymétrique, tandis que la formation du spermatozoïde est symétrique[4]. Pour un mâle, la méiose commence par une cellule diploïde et se termine par quatre spermatozoïdes[5]. Chez les mammifères, le pronucléus femelle commence au centre de l'œuf avant la fécondation. Lorsque le pronoyau mâle est formé, une fois que le spermatozoïde a atteint l'ovule, les deux pronoyaux migrent l'un vers l'autre. Cependant, chez l'algue brune Pelvetia, le pronucléus de l'œuf commence au centre de l'œuf avant la fécondation et reste au centre après la fécondation. En effet, dans les ovules de l'algue brune Pelvetia, le pronucléus de l'œuf est ancré par des microtubules, de sorte que seul le pronucléus mâle migre vers le pronucléus femelle[6].
Concentration de calcium
La concentration de calcium dans le cytoplasme de l'ovule joue un rôle très important dans la formation d'un œuf femelle activé. S'il n'y a pas d'influx de calcium, la cellule diploïde femelle produira trois pronucléi au lieu d'un seul. Cela est dû à l'échec de la libération du deuxième corps polaire[7].
Combinaison de pronoyaux mâles et femelles
Pour les oursins, la formation du zygote commence par la fusion des noyaux interne et externe des pronucléi mâle et femelle. On ne sait pas si l'un des pronoyaux commence la combinaison des deux, ou si les microtubules qui aident à la dissolution des membranes commencent l'action[8]. Les microtubules qui réunissent les deux pronoyaux proviennent du centrosome du spermatozoïde. Il existe une étude qui soutient fortement que les microtubules ont un rôle important dans la fusion des pronoyaux. La vinblastine est un médicament chimiothérapeutique qui affecte à la fois les extrémités positives et négatives des microtubules[9]. Lorsque la vinblastine est ajoutée à l'ovule, le taux d'échec de la fusion des pronuclei est élevé. Ce taux élevé d'échec de la fusion des pronuclei suggère fortement que les microtubules jouent un rôle majeur dans la fusion du pronucléus[10]. Chez les mammifères, les pronuclei ne durent dans la cellule qu'environ douze heures, en raison de la fusion du matériel génétique des deux pronuclei au sein de l'ovule. De nombreuses études sur les pronucléi ont porté sur les ovules d'oursins, où les pronucléi sont dans l'ovule pendant moins d'une heure. La principale différence entre le processus de fusion du matériel génétique chez les mammifères et les oursins est que chez les oursins, les pronucléi entrent directement dans la formation d'un noyau zygote. Dans les ovules de mammifères, la chromatine des pronucléi forme des chromosomes qui fusionnent sur le même fuseau mitotique. Le noyau diploïde chez les mammifères est d'abord vu au stade deux cellules, alors que chez les oursins, il se trouve d'abord au stade zygote[3].
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Pronucleus » (voir la liste des auteurs).
- « Incidence and development of zygotes exhibiting abnormal pronuclear disposition after identification of two pronuclei at the fertilization check », Fertil. Steril., vol. 94, no 3, , p. 965–970 (PMID 19476942, DOI 10.1016/j.fertnstert.2009.04.018)
- Hamoir, « The discovery of meiosis by E. Van Beneden, a breakthrough in the morphological phase of heredity. », The International Journal of Developmental Biology, vol. 36, no 1, , p. 9–15 (PMID 1627480, lire en ligne)
- Scott F. Gilbert et Scott F. Gilbert, Developmental Biology, Sinauer Associates, (ISBN 978-0-87893-243-6, lire en ligne)
- Schmerler et Wessel, « Polar bodies-more a lack of understanding than a lack of respect », Molecular Reproduction and Development, vol. 78, no 1, , p. 3–8 (PMID 21268179, PMCID 3164815, DOI 10.1002/mrd.21266)
- Gorelick, « Meiosis Is Not Gender Neutral », BioScience, vol. 62, no 7, , p. 623–624 (DOI 10.1525/bio.2012.62.7.2, S2CID 85746751)
- Swope et Kropf, « Pronuclear Positioning and Migration during Fertilization in Pelvetia », Developmental Biology, vol. 157, no 1, , p. 269–276 (PMID 8482416, DOI 10.1006/dbio.1993.1131)
- Miao, Stein, Jefferson et Padilla-Banks, « Calcium influx-mediated signaling is required for complete mouse egg activation », Proceedings of the National Academy of Sciences of the United States of America, vol. 109, no 11, , p. 4169–4174 (PMID 22371584, PMCID 3306664, DOI 10.1073/pnas.1112333109, JSTOR 41507114, Bibcode 2012PNAS..109.4169M)
- Longo et Anderson, « The Fine Structure of Pronuclear Development and Fusion in the Sea Urchin, Arbacia punctulata », The Journal of Cell Biology, vol. 39, no 2, , p. 339–368 (PMID 5677969, PMCID 2107533, DOI 10.1083/jcb.39.2.339, JSTOR 1605485)
- Panda, Jordan, Chu et Wilson, « Differential Effects of Vinblastine on Polymerization and Dynamics at Opposite Microtubule Ends », Journal of Biological Chemistry, vol. 271, no 47, , p. 29807–29812 (PMID 8939919, DOI 10.1074/jbc.271.47.29807)
- Hamilton, Suhr-Jessen et Orias, « Pronuclear fusion failure: an alternate conjugational pathway in Tetrahymena thermophila, induced by vinblastine. », Genetics, vol. 118, no 4, , p. 627–636 (PMID 3366365, PMCID 1203318, DOI 10.1093/genetics/118.4.627)