Organe de Haller
L'organe de Haller est une paire d'organe sensoriel possédé par les tiques « dures »[1] (Ixodes ricinus par exemple), décrit pour la première fois[2] par Haller en 1881 (sur un Ixode).
.jpg.webp)
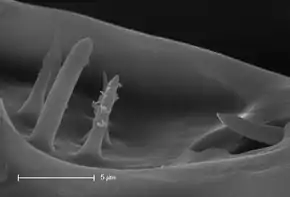
Localisation
L'organe de Haller est situé dans une protubérance près de l'extrémité du dernier article (le tarse) de la première paire de pattes[3]. Cette protubérance est située sur la zone externe (dorsale) du tarse[4].
Description
Cet organe est un « complexe sensoriel » [5]constitué de :
- une capsule fermée dite capsule proximate.
- d'une petite cupule antérieure, formant un minuscule cratère directement ouvert sur la capsule et en contact avec l'atmosphère qui baigne l'environnement de la tique.
- diverses structures qui évoquent des poils ou des épines ; présentes dans les deux cavités il s'agit de sensilles (soies mécanosensibles et chémosensorielles. Selon les espèces, chez l'adulte elles sont au nombre de 15 à 20 (dont seules 6 ou 7 sont visibles dans la dépression ouverte sur l'extérieur).
Quelques-unes de ces soies sont garnies de pores plus ou moins fins selon la soie considérée, et elles sont parfois ramifiées (dendritiques). Toutes sont multi-innervées (connectées à plusieurs neurones). Divers auteurs (tels Lees en 1948[6] puis Arthur en 1962[7] et beaucoup d'autres) estiment que ce soies sont sensibles aux conditions thermohygrométriques et aux odeurs (molécules volatiles émises par les proies de la tique)[8], ce qui ferait de cet organe un organe chémorécepteur. L'étude des sensibles montre que leur lumière (tube intérieur) est associé à des muscles et à des bouquets de fibres nerveuses.
La forme de l'organe de Haller varie beaucoup selon l'espèce, et aussi parfois entre mâle et femelle de la même espèce. Par exemple l'ouverture en est généralement réduite, mais chez les tiques femelles d'un genre spécialisé dans le parasitage de microchiroptères elle est au contraire très large[9].
Fonctions
L'organe de Haller permet aux tiques de détecter la vapeur d'eau[10] (dont elles ont besoin et que certaines espèces peuvent absorber, selon les travaux de Gaede & Knülle publiés en 1997[11]) et le contact avec l'eau liquide (que les tiques préfèrent éviter, même si elles recherchent généralement une hygrométrie importante[12] - [13]) ou la présence d'un animal et notamment des mammifères, par chimiotactisme ou hydrotactisme sur des distances atteignant 10-15m (plus ou moins selon les conditions de circulation de l'air et thermohygrométrique) selon les résultats des travaux de Leonovich publiés en 2004[14].
Cet organe remplirait donc des fonctions comparables à celle des organes olfactifs et gustatifs des mammifères. Et en écartant ses pattes antérieures (à la manière d'un insectes disposant d'antennes), la tique peut probablement détecter des différences entre les flux de molécules analysés par chaque patte, et en déduire la direction d'une source de molécules d'intérêt pour elle.
L'organe de Haller complète les fonctions d'autres organes sensoriels (cellules photosensibles rudimentaires (chez certaines espèces uniquement, les autres étant réputées aveugles au spectre lumineux), pédipalpes, soies distribuées sur les pattes et les téguments du reste du corps).
Outre la vapeur d'eau, et les marques de leurs propres excréments (une expérience simple montre que la tique I. ricinus fait demi-tour quand elle arrive sur une bande de papier « contaminée » par ses propres excréments, mais aussi quand elle s'approche de ces excréments sans même un contact)[15], les tiques seraient ainsi en mesure de détecter le dioxyde de carbone, l'ammoniac, le sulfure d'hydrogène et une variété de composés organiques tels que les benzaldéhydes émis par la respiration, l'haleine ou la sueur d'hôtes potentiels. Différents organes récepteurs ciblent des molécules différentes, avec des sensibilités différentes. Des récepteurs reconnaissent les phénols de composés phénoliques tels que le o-chlorophénol, l'o-bromophénol, l'o-méthylphénol et leurs dérivés ; des récepteurs de lactones reconnaissent la γ-valérolactone[14].
Des sensilles à sensibilité butyrique existeraient mais semblent moins importantes qu'on ne l'a d'abord pensé (avant les années 1950).
On a montré chez Amblyomma variegatum, une tique qui infeste volontiers le bétail, cet organe est aussi le récepteur d'une phéromone d'agrégation (quand les mâles ont trouvé hôte, ils y attirent les femelles ainsi que le ferait un «éclaireur». L'un des composants de cette phéromone est le 2-nitrophénol[14] - [16]. L'organe de Haller joue aussi un rôle dans les réponses intraspécifiques d’agrégation et d'accouplement chez d'autres espèces[15].
Enjeux pour la lutte antivectorielle
Alors qu'un grand nombre d'espèces d'insectes sont en régression, le nombre des tiques augmente de manière d'autant plus préoccupante qu'elle est classée comme le second vecteur de maladies infectieuses (derrière le moustique). On cherche donc à développer des répulsifs moins toxiques ou moins écotoxiques que ceux qui sont sur le marché.
Chez la tique, le meilleur répulsif serait celui qui perturberait (uniquement chez les espèces à risque, idéalement), le comportement de recherche des hôtes. Or c'est l’organe de Haller qui a priori permet à la tique (nymphe et adulte) de détecter les hôtes qui lui fourniront son repas de chair et de sang, grâce aux odeurs, au dioxyde de carbone et à la chaleur qu'ils émettent[17] - [18] - [19] - [20] - [21]. En 2018, les mécanismes de perturbation du comportement (de recherche de l'hôte) par les répulsifs ne sont pas encore clairement compris[22] - [23].
Un autre objectif pourrait être de pouvoir sélectivement piéger les tiques, ou de pouvoir les piéger dans certains environnements (jardins...). C'est par l'organe de Halleur qu'elles détecteraient le cocktail chimique (encore à découvrir) qui les conduit habituellement leurs hôtes.
- Thermotaxie : Le rôle du rayonnement infrarouge émis par l'hôte n'était au début des années 2000 pas encore consensuel, car la capacité de l'organe de Haller à le détecter n'était pas scientifiquement établie ou expliquée ; de premières études avaient même conclu que les tiques n’utilisent pas la chaleur radiante. Mais on a récemment ;
- en 2017 D Michel et ses collègues montrent enfin que la tique Dermacentor variabilis adulte est bien attirée par le spectre infrarouge, et que ce phototactisme IR cesse quand les organes de Haller sont supprimés. Lors de cette expérience les tiques étaient aussi attirées par une lumière blanche (avec et sans les organes de Haller, mais uniquement quand leurs ocelles (yeux très primitifs) n'étaient pas obstrués). Si les scelles sont obstruées, la tique ne réagit plus au spectre visible de la lumière, reste attirée par l'infrarouge. Un récepteur putatif de la protéine TRPA1 (Transient receptor potential cation channel, subfamily A, member 1) a été caractérisé à partir d'un transcriptome spécifiquement réalisé pour l'organe de Haller de D. variabilis. Fait intéressant ; ce récepteur était homologue d'un récepteur connu chez les Vipères, les Pythons et Boas (seul récepteur connu en 2017) pour la détection infrarouge chez l'animal.
De nouvelles structures associées à la fosse antérieure (AP) ou à la sensille saillante et à la capsule postérieure de l'organe de Haller ont été décrites, dont certaines pourraient jouer un rôle dans la détection infrarouge [24], - en 2019, on a confirmé [21] la thermotaxie chez Dermacentor variabilis, mais aussi chez Amblyomma americanum que ce type de tiques détectent un être humain à plusieurs mètres de distance grâce au rayonnement thermique qu'il dégage[21].
Le mécanisme commence à être compris : la « capsule » (zone sphérique à noyau arrondi et couvert) située sur l'organe de Haller est munie d'une petite ouverture, qui en fait un détecteur directionnel. L'intérieur de cette capsule est hautement réfléchissant, ce qui lui confère une haute sensibilité[21]. Même en petite quantité, des répulsifs d'insectes ou d'acariens tels que le DEET, la picaridine, la 2-undécanone, le citronellal et le nootkatone éliminent la thermotaxie[21]. Mais elles n'affectent pas la capacité de recherche d’hôte stimulée par les odeurs. De répulsifs plus performants sont donc encore à mettre au point contre les tiques et autres ectoparasites afin de limiter l'exposition des humains et de leurs animaux de compagnie et d'élevage pour le protéger des maladies à transmission vectorielle[21].
- en 2017 D Michel et ses collègues montrent enfin que la tique Dermacentor variabilis adulte est bien attirée par le spectre infrarouge, et que ce phototactisme IR cesse quand les organes de Haller sont supprimés. Lors de cette expérience les tiques étaient aussi attirées par une lumière blanche (avec et sans les organes de Haller, mais uniquement quand leurs ocelles (yeux très primitifs) n'étaient pas obstrués). Si les scelles sont obstruées, la tique ne réagit plus au spectre visible de la lumière, reste attirée par l'infrarouge. Un récepteur putatif de la protéine TRPA1 (Transient receptor potential cation channel, subfamily A, member 1) a été caractérisé à partir d'un transcriptome spécifiquement réalisé pour l'organe de Haller de D. variabilis. Fait intéressant ; ce récepteur était homologue d'un récepteur connu chez les Vipères, les Pythons et Boas (seul récepteur connu en 2017) pour la détection infrarouge chez l'animal.
Notes et références
- « Organe de Haller - Dictionnaire des Sciences Animales », sur cirad.fr (consulté le ).
- Haller G (1881) Vorliiufige Bemerkungen iiber das Geh6rorgan der Ixodiden. Zool. Anz. 4, 165-167
- « Pfizer.ch », sur Pfizer.ch (consulté le ).
- Parola P & Raoult D (2001) Ticks and tickborne bacte rial diseases in humans: an emerging infectious threat. Clinical infectious diseases 32:897-928.
- Schramm F (2012). Inflammation cutanée et borréliose de Lyme: étude in vitro des interactions entre les cellules résidentes de la peau et Borrelia (Thèse de Doctorat, Université de Strasbourg)
- Lees A.D (1948) The sensory physiology of the sheep tick, Ixodes ricinus. J. Exp. Biol. 25 , 145–207
- Arthur Don R (1962) Ticks and disease. Pergamon press, intern. series of monogr. on pure and applied Biol. ZOO~. Div., 9.
- Waladde S.M (1982) '[Tip-recording from ixodid tick olfactory sensilla: responses to tick related odours. Journal of comparative physiology, 148(4), 411-418 |résumé.
- Morel, P. C., & Perez, C. (1973). Morphologie des stades préimaginaux des Ixodida s. str. d’Europe occidentale. II.-Les larves des Pholeoixobh Schulze, 1942.
- Hair J.A, Sauer J.R & Durham K.A (1975) Water balance and humidity preference in three species of ticks. J. Med. Ent. 12 , 37–47
- Gaede K and Knülle W (1997) On the mechanism of water vapour sorption from unsaturated atmospheres by ticks. J. Exp. Biol. 200 , 1491–1498
- Krober, T., & Guerin, P. M. (1999). Ixodid ticks avoid contact with liquid water. Journal of Experimental Biology, 202(14), 1877-1883 (PDF, 7 pages).
- Fielden L.J & Lighton J.R.B (1996) Effects of water stress and relative humidity on ventilation in the tick Dermacentor andersoni (Acari: Ixodidae). Physiol. Zool. 69 , 599–617
- Leonovich S.A. (2004) Phenol and lactone receptors in the distal sensilla of the Haller’s organ in Ixodes ricinus ticks and their possible role in host perception. In: Exp Appl Acarol ;32(1-2), S. 89-102,
- Grenacher S, Kröber T, Guerin P.M & Vlimant M (2001) Behavioural and chemoreceptor cell responses of the tick, Ixodes ricinus, to its own faeces and faecal constituents. Experimental & applied acarology, 25(8), 641-660 (résumé).
- Diehl P.A, Guerin P.M, Vlimant M &Steullet P (1991) Biosynthesis, production site and emission rates of aggregation-attachment pheromone in males of two Amblyomma ticks. J. Chem. Ecol. 17 , 833–847
- LEES, A. D. The Sensory Physiology of the Sheep Tick, Ixodes Ricinus L. Journal of Experimental Biology 25, 145-207 (1948).
- Apanaskevich, D. A. & Oliver, J. H. J. in Biology of TIcks Vol. 1 (eds D. E. Sonenshine & R. M. Roe) Ch. 3, 59-73 (Oxford University Press, 2014).
- Leonovich, S. A. Orientatinal Behavior of the Ixodid Tick Hyalomma Asiaticum Under Desert Conditions. Parazitologia 20, 431-440 (1986).
- Carr, A. L. & Roe, M. Acarine attractants: Chemoreception, bioassay, chemistry and control. Pestic Biochem Physiol 131, 60-79, doi:10.1016/j.pestbp.2015.12.009 (2016)
- Carr A.L & Salgado V (2019) Ticks Home in on Body Heat: A New Understanding of Ectoparasite Host-Seeking and Repellent Action. bioRxiv, 564179.
- Wikel, S. K. Ticks and Tick-Borne Infections: Complex Ecology, Agents, and Host Interactions. Vet Sci 5, 60, doi:10.3390/vetsci5020060 (2018)
- Sonenshine, D.E & Roe R.M (2014) Biology of Ticks. Vol. II (Oxford University Press).
- Robert Mitchel D & al (2017) Infrared light detection by the haller’s organ of adult american dog ticks, Dermacentor variabilis (Ixodida: Ixodidae) ; Volume 8, Issue 5, August 2017, Pages 764-771 ; Ticks and Tick-borne Diseases - Short communication | https://doi.org/10.1016/j.ttbdis.2017.06.001
Voir aussi
Articles connexes
Lien externe
- Photographie de profil d'un Organe de Haller (xodes ricinus) en microscopie optique, par Paul Leroy | Licence CC BY NC, montrant la fosse recouverte d'une fine pellicule translucide
{kind=link}
Bibliographie
- Les tiques : identification, biologie, importance médicale et vétérinaire, Ed. Lavoisier, 2007 (ISBN 9782743019563)
- Grand dictionnaire illustré de parasitologie médicale et vétérinaire
- Bruce W.A (1971) Posterior capsule of Haller's organ in the lone star tick Amblyomma americanum (Acari: Ixodidae). The Florida Entomol. 64, 65-72.
- Dautel, H. and Knülle, W.(1997). Cold hardiness, supercoolingability and causes of low-temperature mortality in the soft tick, Argas reflexusand the hard tick, Ixodes ricinus(Acari, Ixodoidea)from Central Europe. J. Insect Physiol.43, 843–854
- Elizarov Y.A (1963) Insect and tick chemoreception: Electrophysiologic study of the chemoreception of Ixodidae. Vestn. Mosk. Univ. Ser. VI Biol. Pocvoved. 16 , 16–24
- Estrada-Pena, A., Lucientes-Curdi, J., Gutierrez-Galindo, J. F., Sanchez-Acedo, C., Ocabo-Melendez, B., Castillo-Hernandez, J. A., & Galmes-Femenias, M. (1986). Morfologia del órgano de Haller en garrapatas (Ixodoidea). Revista iberica de parasitología, 46(2), 175-179.
- Foelix R.F & Axtell RT (1972) Ultrastructure of Haller's Organ in the Tick Amblyomma americanum. In: Zeitschrift für Zellforschung. 124, S. 275-292
- Foelix R.F & Axtell RT (1971) Fine structure of tarsal sensilla in the tick Amblyomma americanum (L.). Z. Zellforsch. 114, 22-37 (1971)
- Hess E & Vlimant M (1986) Leg sense organs of ticks. In Morphology, Physiology and Behavioural Biology of Ticks (ed. J.R Sauer & J.A Hair), pp. 361–390. Chichester : Ellis Horwood.
- Jorgensen W.K (1984). The ultrastructure of the sense organ of tarsus I, the palps and chelicerae of the larval cattle tick Boophilus microplus (Canestrini) Ixodidae, including a study of the ultrastructure and function of the cattle tick podium. PhD thesis, University of Queensland, Brisbane, Australia. 236pp.
- Krober T & Guerin P.M (1999) Ixodid ticks avoid contact with liquid water. Journal of Experimental Biology, 202(14), 1877-1883.
- Leonovich S.A. (2004) Phenol and lactone receptors in the distal sensilla of the Haller’s organ in Ixodes ricinus ticks and their possible role in host perception. In: Exp Appl Acarol ;32(1-2), S. 89-102,
- Nuttall G., Cooper W & L Robinson L (1908). "On the structure of "Haller's organ" in the Ixodoidea". Parasitology 1 (3): 238–242. doi:10.1017/S0031182000003486
- Steullet P & Guerin P.M (1994). Identification of vertebrate volatiles stimulating olfactory receptors on tarsus I of the tick Amblyomma variegatum Fabricius (Ixodidae). I. Receptors within the Haller’s organ capsule. J. Comp. Physiol. A 174 , 27–38
- Steullet P (1993) Perception of vertebrate volatiles in the tropical bont tick, Amblyomma variegatum Fabricius. Thèse présentée à la Faculté des Sciences de l'Université de Neuchâtel pour obtenir le grade de docteur es sciences.
- Thonney F (1987) Étude morphologique et structurale des récepteurs sensoriels du tarse I de la tique Ixodes ricinus L. Thèse de doctorat, Université de Neuchâtel, Neuchâtel. 135pp
- Zolotarev E.Kh & Elizarov Yu. A (1963) Studies of chemoreception in insects and ticks. Location of the chemoreceptors in the tick Ixodes persulcatus P. Sch. Vestn. Mosk. Univ. 18, 7-9 .
- Zolotarev E.Kh & Elizarov Yu. A (1964) Investigation of the chemoreception in insects and ticks: The pecularities of the functioning of chemoreceptors in Hyalomma asiaticum P. Sch. et E. Sch. under the action of repellents. Zoo!. J. Mosk. 43, 549-559
- Zolotarev E.Kh & Sinitsyna Y.Y (1965) Chemoreceptive organs on the fore legs of ixodid ticks. Vestn. Mosk. Univ. 20, 17-2.
- Леонович С.А (2004) Тонкое строение тарзальных сенсилл таежного клеща Ixodes persulcatus (Ixodidae) (La structure subtile des sensilles du tarse de la tique de la taïga Ixodes persulcatus (Ixodidae)). Паразитология, (4), 304-309 (ru).