Nouveauté évolutive
Une nouveauté évolutive est quelque chose, une structure ou une fonction par exemple, qui semble qualitativement distincte de ce qui était présent auparavant chez un organisme.

-eye.JPG.webp)
Cependant, il s'agit d'un concept relativement nouveau, qui s'apparente au domaine de recherche en biologie évolutive du développement. À ce jour, il n'y a pas de consensus quant à la définition précise des nouveautés évolutives[1].
La carapace des tortues, les yeux, les cornes des scarabées et les membres des tétrapodes sont quelques exemples de nouveauté peu contestés[2] - [3].
Nouveauté ou modification?
La recherche sur les nouveautés évolutives a pour but d'expliquer leur origine en s'intéressant aux modifications du plan corporel à différents niveaux d'analyse[4]. Ce dernier facteur est déterminant quand il est question de départager ce qui est nouveau de ce qui est modifié. Il existe une vaste gamme de nuances entre une modification et une nouveauté et le positionnement d'une observation quelque part dans ce gradient est intrinsèquement dépendant de l'analyse qui en est fait. Par exemple, ce qui peut sembler nouveau à la suite d'une analyse fossile semblera n'être guère plus qu'une modification à la suite d'une analyse cytologique[2]. Il s'agit donc d'un concept empreint de subjectivité, car plus le «grain» d'analyse est fin, plus on a de chances de trouver une explication[5]. Mais jusqu'à quel degré une explication discrimine une nouveauté d'un simple changement? Encore ici, il n'y a pas d'entente à ce sujet.
Homologie profonde et nouveauté
On distingue généralement aisément une nouveauté d'une homologie primaire (au niveau morphologique), mais cette distinction n'est plus nette dès qu'on parle de génétique. Les homologies profondes permettent de constater qu'une immense quantité de formes, de traits et de fonctions sont réalisables simplement par la combinaison d'un nombre restreint d'outils[3]. C'est donc la nouveauté de la combinaison, génératrice d'un nouveau résultat, qui est importante.
Les GRNs
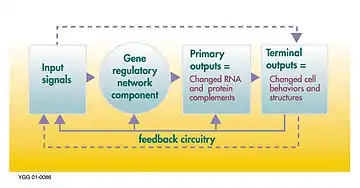
Les réseaux de régulation génétique (GRN) sont les boîtes à outils des nouveautés. Ce sont eux qui gèrent les patrons de développement et c'est majoritairement sur la différence dans leur régulation ainsi que leur composition que se base la divergence dans les plans corporels[6]. Il est donc primordial de connaître les GRNs, de connaître leur composantes, la ou les fonctions de chacune d'entre elles, et de savoir ce qui les lies entre elles pour comprendre les nouveautés évolutives[4]. Le moment dans le développement auquel se produit le changement, ainsi que sa magnitude, c'est-à-dire l'ampleur de ses répercussions, sont déterminants quant à la viabilité de la nouveauté ainsi engendrée[7] - [8].
Contraintes à l'apparition de nouveautés
De nombreux effets pléiotropiques négatifs peuvent résulter d'une ou de plusieurs modifications dans un GRN[7]. Considérant la grande connectivité de ses composantes, un infime changement peut avoir de vastes répercussions indésirables sur les structures environnantes.
Plus ces phénomènes se produisent tôt dans le développement, plus leur impact est grand, plus le biais de développement sera important et négatif et moins ils aura de change qu'un nouveauté évolutive émerge dans une population[8].Tous les stades du développement qui précèdent l'organogenèse sont sous un contrôle beaucoup plus stricte que ceux qui succèdent ce stade.
De plus, la force de la sélection interne peut être plus grande selon les taxas. Par exemple, les invertébrés présentent un développement plus plastique que les vertébrés[6], c'est-à-dire qu'ils sont soumis à moins de contraintes lorsqu'un changement survient dans leur patron de développement.
Ce ne sont pas toutes les nouveautés qui réussissent à voir le jour, et donc qui ont la change d'être testée par la sélection naturelle. Par exemple, les mutations homéotiques peuvent parfois être si drastiques que le développement n'arrive à terme que dans peu de cas. C'est ce qu'on observe notamment au niveau du nombre de vertèbres cervicales chez l'humain[9]. Les effets pléiotropiques négatifs qui y sont associés sont très forts, car il s'agit d'une modification qui survient très tôt dans le développement et le taux de survie est pratiquement nul.
Sources de nouveautés
Il existe plusieurs phénomènes qui peuvent être considérés comme des sources de nouveautés, car ils agissent sur les GRNs en en modifient la composition ou le fonctionnement. N'importe quel de ces changements aura une ou des répercussions, directes ou indirectes, sur le plan corporel d'un organisme. C'est le degré d'impact de ces modifications qui détermine s'il sera facile ou non pour la nouveauté de s'implanter.
Modification de l'ADN
Les caractéristiques génétiques qui sous-tendent à une nouveauté peuvent théoriquement toujours être retrouvées. Le plus souvent, c'est la modification des régions régulatrices en amont des régions codantes qui mène à l'apparition de fonctions ou caractères nouveaux[10]. Puisque les gènes ont souvent plusieurs fonctions, l'organisme est capable de les réguler indépendamment par des promoteurs différents. C'est pourquoi une mutation dans une région régulatrice peut ne pas affecter toutes les fonctions prises en charge par un même gène. La spécificité de cette régulation est favorable à l'apparition de nouveautés, car plus un changement affecte un grand nombre de composantes du patron du développement, plus il y a de chances que des effets négatifs y soient liés.
Même si elles sont moins susceptibles d'être positives, les mutations dans les régions codantes peuvent offrir un potentiel évolutif pour les nouveautés[10]. Si une séquence codant une protéine est altérée, la protéine résultante aura une forme différente, donc une fonction différente. Cette protéine peut elle-même être un facteur de transcription, qui ira affecter l'expression de d'autres gènes dans un GRN.
Duplication
La duplication des gènes est probablement la source de nouveautés la plus importante[7]. Elle offre un grand potentiel évolutif, car un gène présent en deux copies représente une redondance, c'est-à-dire qu'il a plus de matériel que nécessaire pour l'accomplissement d'une même fonction. La duplication offre donc une opportunité de néofonctionnalisation et d'altération du patron d'expression génétique[11].Ces gènes viennent avec tous les éléments clés pour bien fonctionner et c'est la ou les modifications qu'ils subirons qui leur permettra de donner naissance à de nouvelles structures ou fonctions.
La duplication de groupes de gènes Hox en entier est un phénomène majeur dans l'émergence des nouveautés[6] - [10]. Il est étonnant de constater que, chez des organismes très différents morphologiquement, ce sont souvent des groupes de gènes Hox très similaires qui sont responsables du développement des mêmes structures[10]. Leur duplication offre donc un grand potentiel évolutif.
Co-option
Il est aussi important de considérer la co-option dans l'apparition de nouveautés[10]. Ces dernières ne sont pas causées que par des changements dans le fonctionnement des GRNs, mais aussi par des ajouts de fonctions. Un ajout de fonction change la dynamique du patron de développement et de nouvelles options deviennent alors disponibles.
Un environnement propice
Les organismes présentant des nouveautés qui réussissent à passer à travers la sélection interne ne sont pas encore arrachées à leur misère. Il s'ensuit la sélection naturelle, qui dépend de beaucoup de facteurs. Le ou les individus portant une nouveauté doivent être capable de la transmettre efficacement afin qu'elle persiste dans une population et éventuellement chez tous les individus de l'espèce.
La fréquence des apparitions des nouveautés peut être liée à l'environnement qui les entoure. Plus un événement qui interfère dans le patron du développement arrive souvent dans un individu, plus il y a de chances qu'éventuellement, cette modification puisse être intégrée et engendre une nouveauté au sein de l'organisme et de la population. Cette idée prend tout son sens lorsqu'on s'intéresse aux nouveautés évolutives d'un point de vue géographique et paléontologique[12]. On suppose que la compétition élevée pour les ressources et les niches écologiques aux basses latitudes aurait favorisé l'apparition de formes diverses et nouvelles avec une faible répression. Cet environnement serait donc propice à l'expérimentation, car il devient plus rentable de changer que de rester stable. Les tropiques seraient donc en quelque sorte «un accumulateur de diversité»[12].
Cette course évolutive peut être liée à des périodes de relâchement de la sélection, qui peuvent augmenter les chances d'une nouveauté de persister. La colonisation de nouveaux habitats, la diminution de la prédation ou l'augmentation des ressources en sont trois exemple[6] - [7]. Si un changement important dans le patron du développement coïncide avec le début d'une radiation adaptative, il est possible qu'une nouveauté s'avère pratique pour une espèce sous de nouvelles conditions. Le lien entre les radiations adaptatives et les nouveautés évolutive ne fait pas l'unanimité dans la communauté scientifique. Une radiation adaptative sans l'apparition de nouveautés à proprement parler est possible, mais est-ce qu'une nouveauté peut apparaître sans un radiation? Il n'y a pas de réponse claire à cette question.
La carapace des tortues
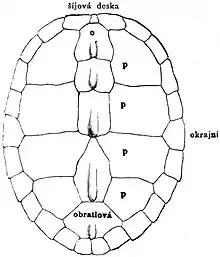

Un simple coup d'œil sur une carapace de tortue est suffisent pour émettre l'hypothèse qu'il s'agit d'une nouveauté évolutive. Il est difficile de percevoir le lien qui existe entre les chéloniens et les autres vertébrés[13]. Alors comment une structure aussi unique a-t-elle pu se former?
Il faut d'abord souligner les différences majeurs entre le plan corporel de la tortue et celui des autres vertébrés, qui est le résultat d'un changement dans le patron du développement du squelette des tortues[13]. Alors que les omoplates des vertébrés se retrouvent à l'extérieur de la cage thoracique, celles de la tortue sont plutôt à l'intérieur[14]. Ce phénomène a également un impact sur le point d'attachement de deux muscles : le muscle dentelé antérieur et le muscle élévateur de la scapula. Les muscles chez la tortue s'attachent dorsalement et ventralement à l'intérieur de la cage thoracique, alors qu'ils sont externes chez les autres vertébrés. Le changement dans la position de l'omoplate et dans l'attachement des muscles est causé par la croissance particulière des côtes.
La carapace des tortues provient de l'acquisition de l'ostéoderme par les chéloniens. C'est l'élargissement des côtes et des vertèbres, liés à l'ostéoderme, qui forme cette structure. Une des hypothèses les plus probable soutient qu'un phénomène d'hétérotopie pourrait être à la base de la formation de la carapace[15]. Le développement osseux aurait changé de région, ce qui aurait modifié la nature des interactions «entre le derme et le primordium des côtes, menant à une ossification dermale ectopique»[13].
La croissance particulière des côtes est causée pas les primordium des crêtes carapaciales, des structures au niveau tissulaire spécifique aux tortues[13]. Ces crêtes seraient dérivées des crêtes wolfiennes des amniotes, impliquées notamment dans la formation du système urogénital et dans le développement des membres. Communément appelées CR, les crêtes carapaciales sont à la limite entre le plan axial et le plan latéral du corps de la tortue. Ce sont elles qui influencent la croissance des côtes selon un modèle en plaques, qui donne naissance à la carapace.
Plusieurs gènes présents dans les CR seraient dévirés de gènes qu'on retrouve chez les amniotes[13]. L'altération de leur expression aurait mené à la formation des CR. Il s'agit donc d'un exemple de co-option, puisque ces gènes possèdent également d'autres fonctions.
Le rôle des CR dans l'arrêt de la croissance des côtes vers l'axe latéral du corps ainsi que dans l'internalisation de certaines structures est encore peu connu, mais il serait plutôt indirecte[13] - [14]. Durant le développement, «la seconde côte, la plus postérieure, croisse latéralement et antérieurement par-dessus l'omoplate et le muscle dentelé antérieur en pliant la partie dorsale de la paroi latérale du corps vers l'intérieur.»[14]. Les CR sont retrouvées au long de ce plie et ont donc une forte influence sur la forme globale du corps des tortues.
Le seul intermédiaire entre les vertébrés et les chéloniens qui a été trouvé jusqu'à maintenant est Odontochelys semitestacea, un fossile de 220 millions d'années[14] . Cette espèce possède un plastron, des côtes élargies dont la croissance latérale est restreinte, mais pas de carapace ni d'internalisation de l'omoplate. Cette dernière se retrouve plutôt complètement au devant les côtes. Ces différences serait potentiellement causée par l'absence de CR antérieurs et postérieurs et ce n'est que lorsqu'ils entourent complètement l'animal que la carapace peut se former. Malgré la présence dans les fossiles de cet intermédiaire, le développement corporel des tortues reste très singulier et cette différenciation serait le résultat d'un important biais de développement, plutôt que d'une accumulation de petits changement à travers un grand nombre de génération.
En somme, on observe chez les chéloniens une modification du plan corporel par une combinaison d'hétérotopie, de co-option et d'autres phénomènes au niveau moléculaire encore peu connus. Ici, une nouveauté est donc considéré comme une combinaisons nouvelle d'éléments préexistants qui engendre un changement non gradué menant à la formation d'une nouvelle structure.
Voir aussi
Articles connexes
Notes et références
- (en) 2010, « Evolutionary Novelties and the Evo-Devo Synthesis : Field Notes », sur University of Alberta (consulté le ).
- Pigliucci, M., « What, if anything, is an evolutionary novelty? », Philosophy of Science,vol.75, n⁰5, 2008, p.887-898
- Shubin, N., « Deep homology and the origins of evolutionary novelty », Nature, vol.457, n⁰7231, 2009, p.818-823
- Wagner, G. P., Lynch, V. J.,« Evolutionary novelties », Current Biology, vol.20, n⁰2, 2010, p.48-52.
- (en) Wilkins, J. S.,, « Notes on Novelties », sur Evolving Thoughts, 2011-2012 (consulté le )
- Wagner, G. P., Amemiya, C., Ruddle, F., « Hox cluster duplications and the opportunity for evolutionary novelties », Proceedings of the National Academy of Sciences, vol.100, n⁰25, 2003, p.14603-14606.
- Galis,F., Metz, J. A.,« Evolutionary novelties : the making and breaking of pleiotropic constraints », Integrative and comparative biology, vol.47, n⁰3, 2007, p.409-419.
- Arthur, W., « Intraspecific variation in developmental characters: the origin of evolutionary novelties », American Zoologist, vol. 40, n⁰3, 2000, p. 409-419.
- Galis, F., Van Dooren, T. J., Feuth, J. D., Metz, J. A., Witkam, A., Ruinard, S., Wunaendts, L. C., « Extreme selection in humans against homeotic transformations of cervical vertebrae », Evolution, vol.60, n⁰12, 2006, p.2643-2654.
- Pick, L., Heffer, A., « Hox gene evolution: multiple mechanisms contributing to evolutionary novelties », Annals of the New York Academy of Sciences, vol.1256, n⁰1, 2012, p.15-32.
- Lynch, M., Conery, J. S., « The evolutionary fate and consequences of duplicate genes », Science, vol.290, n⁰5494, 2000, p. 1151-1155.
- Jablonski, D., « The tropics as a source of evolutionary novelty through geological time », Nature, vol.364, n⁰6433, 1993, p.142-144.
- Kuratani, S., Kuraku, S., Nagashima, H., « Evolutionary developmental perspective for the origin of turtles: the folding theory for the shell based on the developmental nature of the carapacial ridge », Evolution and Development, vol.13, n⁰1, 2011, p. 1-14.
- Nagashima, H., Sugahara, F., Takechi, M., Ericsson, R. M., Kawashima-Ohya, Y., Narita, Y., Kuratani, S., « Evolution of the turtle body plan by the folding and creation of new muscle connections », Science, vol.325, n⁰5937, 2009, p. 193-196.
- Gilbert, S. F., Cebra-Thomas, J. A., Burke, A. C., « How the turtle gets irs shell », Biology of Turtles, CRC Press, Boca Raton In J. Wyneken, M. H Godfrey, and V. Bels (eds.), 2008, p. 1-16.