Modularité biologique
La modularité biologique est un concept de la biologie évolutive du développement : de petits modules, sous-unités de l’organisme, qui fonctionnent indépendamment les uns des autres, permettent une structure et une logique chez l'organisme fonctionnel, par le biais de la communication intermodule. Ainsi ils interagissent pour former un tout fonctionnel qui peut évoluer et se développer[2]. Ceci vaut aussi à l'échelle d'une métapopulation (constituée d'un réseau de sous-population pouvant être considérée chacune comme un module[3]).
La modularité représente aussi le fait qu'un module (ex. une cellule d'un certain type) se duplique pour former un organisme dont tous les modules sont semblables. Ce sont donc les deux définitions généralement acceptées pour décrire la modularité. Même si ces définitions restent vagues, une seule chose est primordiale pour décrire un module. Ce module doit avoir des interactions internes plus fortes ou plus régulières que ses interactions avec les autres modules qui l'entourent[4].
La modularité est difficile à définir car un module est différent selon le milieu de recherche en biologie. S’il s’agit de chercheurs en évolution, un module sera une partie d’un organisme, tel un organe, tandis que les chercheurs en développement décriront les modules comme étant des composants plus petits, comme des gènes. La définition peut aussi être différente selon la direction de la recherche. Dans une vision expérimentale, un module est un ensemble de structure avec une fonction commune, tandis que dans une vision comparative, il s’agit d’un patron structurel[5].
Pour permettre plus de clarification, une définition complète de la modularité devrait comprendre les éléments suivants :
- Un module est une structure ou un processus qui subit des interactions davantage internes qu’externes ;
- Un module est délimité du milieu, a une fonction qui lui est propre et est donc un individu en soi ;
- Un module est séparé des autres modules avec lesquels il peut interagir ;
- Un module peut intervenir à n’importe quel niveau biologique[5].
Éléments d'explications
La modularité se distingue en plusieurs concepts,
- soit c’est une parcelle d’un individu (ou d'une population) servant au fonctionnement et au maintien de l’organisme viable (ou de la métapopulation) ;
- soit c’est une cause de l’évolution[6].
L’idée de l’organisation modulaire chez les individus, dans un contexte évolutif, a été proposée pour la première fois par M. Riedl en 1978 ; M. Lewontin et M. Bonner ont suivi cette même réflexion peu de temps après, mais en abordant différemment cette perspective. Au début, cette idée n’a pas fait grand bruit dans la communauté scientifique jusqu’aux récentes découvertes allant dans ce sens[7].
John Tyler Bonner introduit l’idée de réseau génique, posant l'hypothèse que les changements pleiotropiques sont un exemple de modularité, car permettant le changement de certains caractères sans affecter les autres caractères. Il affirme en outre que ces changements indépendant, au niveau modulaire, permettent une plus grande viabilité pour l’individu face à ces changements[8]. Les variations modulaires au sein de l'organisme n'interrompent pas le fonctionnement de l'organisme entier. Donc, un module peut modifier ses possibilités phénotypiques sans affecter les autres modules[9].
Exemples
Modularité dans la cellule
La modularité se retrouve au niveau des gènes de régulation cis du développement, section de gènes qui se retrouvent sur le même chromosome et qui entraînent le développement par cascade de molécules activatrices ou inhibitrices, qui entraîne la formation de protéines produites par une cellule. Ces protéines possèdent, elles aussi, un arrangement modulaire[10].
Modularité au niveau protéique
Les protéines sont présentes partout dans l'organisme, mais aussi sont présentes tout au long du développement et bien plus loin, lors de la vie entière de l'organisme. Les protéines possèdent des fonctions diverses. Elles ont un effet sur ce qui est visible de l'organisme, comme le phénotype de l'individu, mais aussi sur ce qui n'est pas visible à l'œil nu, tel que l'acide désoxyribonucléique, la cellule en général et bien plus.
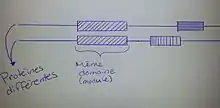
Un petit récapitulatif sur la forme des protéines sera nécessaire pour bien comprendre le concept de module au niveau protéique. Il y a différentes étapes pour atteindre la protéine telle qu'on la connaît. La structure primaire de la protéine est lorsque les acides aminés, la plus petite unité de la protéine, au nombre de 20, s'assemblent ensemble pour former une chaîne linéaire. La structure secondaire est obtenue à la suite du repliement de la structure primaire chaîne d'acide aminé. Cette structure a pour forme finale une hélice alpha ou un feuillet bêta. La structure tertiaire est le résultat du repliement des feuillets bêta ou des hélices alpha. Ceci crée alors une structure en trois dimensions qui est une seule et longue chaîne repliée plusieurs fois. La structure quaternaire est un assemblage de plusieurs chaînes, 2 ou plus, c'est la protéine à sa forme finale. La modularité intervient à ce niveau, car la protéine est composée de modules, structures tertiaires, qui sont indépendantes les uns des autres, mais qui interagissent ensemble pour former une protéine fonctionnelle. Les domaines qui composent la protéine sont des modules, car cette partie se replie indépendamment du reste de la protéine. Toutefois, il est important de noter que l'inverse n'est pas nécessairement applicable[11]. De plus, les structures modulaires d'une protéine, comme le repliement de ces modules de la protéine sont un élément qui est totalement indépendant du milieu dans lequel elles se trouvent[12].
- Deux illustrations
- Tout d'abord, le cas où tous les domaines sont des modules. Certains modules peuvent être homologues, mais la protéine ne l'est pas dans tous les cas. Voici une image pour aider à visualiser ce concept[11].
- Ensuite, un exemple où un module n'est pas un domaine. Deux bons exemples pour cela sont les protéines fibreuses et les structures désordonnées. En effet, l'arrangement de ces protéines permet d'identifier les modules, mais les domaines ne sont pas identifiables ni séparables du reste de la protéine. Un module peut être composé de plusieurs domaines en interactions entre eux. Donc, si ces domaines sont séparés ils ne seront plus fonctionnels, alors ils ne sont pas indépendants[11].
Hétérométrie
Sur le plan physique, la modularité permet la croissance indépendante de différents organes ou appendices du corps de l'organisme, et ce, indépendamment des organes ou appendices adjacents, du même type, et du même individu ou de différentes espèces proches phylogénétiquement. Ces modifications se font à partir des facteurs engendrant ou modifiant le développement de l'individu. La sensibilité aux facteurs de développement peut varier. Il est bon de noter de que de manière générale, c'est une accumulation de plusieurs petits changements phénotypiques qui mène à l'apparition d'adaptation[13].
Modularité chez les animaux
- Chevaux
Les chevaux ancestraux possédaient cinq orteils, dont le médian avait une croissance 1,4 fois plus rapide que les autres ce qui entraînait une augmentation de longueur de cet orteil. Au fil des générations, ces modifications se sont accumulées ce qui a mené aux chevaux actuels à un seul orteil[13].
- Baleine

L'embryon de baleine possède un crâne très semblable à tous les vertébrés, mais c'est plus tard dans le développement que se produisent les transformations. En effet, le développement accéléré des mâchoires supérieures et inférieures mène à un allongement du museau de cet animal. Cet allongement engendre alors le déplacement des fausses nasales vers le dessus de la tête. Ce changement a permis aux baleines de pouvoir respirer tout en gardant son corps parallèle à la surface de l'eau[13].
- Rat kangourou du désert
Les abajoues des rats kangourou ont la même utilité que cette même structure chez les hamsters. Cette structure sert de réserves de nourriture pour ces animaux. Toutefois dans le cas du rat kangourou qui vit en milieu désertique cette structure a une utilité supplémentaire, celle de maintenir les graines récoltées servant à la nutrition dans un environnement humide pour éviter le dessèchement. Cette structure n'a pas évolué exactement de la même manière, car le rat kangourou possède des poils à l'intérieur de ces abajoues, contrairement aux hamsters. De plus, les abajoues des rats kangourou ne communiquent pas avec la cavité orale. Le développement cd cette structure chez les deux organismes est exactement le même au départ, car il prend naissance grâce au repli de l'épithélium de la cavité buccale. Toutefois, la croissance accrue du museau du rat kangourou tire l'ouverture des abajoues vers l'avant à la limite des lèvres et devient éventuellement externe[13].

Crânes de mammifères[14] - [15]
But des études
L’ensemble des mammifères regroupent plusieurs espèces ayant des morphologies crâniales bien différentes. L’équipe de Porto s’est demandé s’il est possible de révéler des modules crâniens similaires à travers la phylogénie des mammifères. Par le fait même, ils se sont demandé si ces modules sont en liens avec une fonction particulière et si la taille du spécimen peut faire une interférence entre le module et la pression de sélection.
Signification des études
Un module est un complexe semi-autonome démontrant un lien entre la fonction et/ou le développement et un trait particulier. Cette structure peut faciliter ou limiter l’évolution d’un trait dans le temps, ce qui rend les modules très importants dans le cadre de recherches en évolution et développement. Le crâne de mammifère est un sujet particulièrement intéressant, puisque les os sont homologues à travers les différents ordres et il prend part à plusieurs fonctions. De plus, comme le patron de développement est relativement stable, il s’agit d’un bon sujet pour une étude comparative.
Méthodes d’expérimentations
Les expérimentations ont été effectuées sur 3644 crânes adultes de 15 ordres de mammifères dans un premier temps et dans un second sur 3641 crânes et 18 ordres. Les crânes ont tous été digitalisés et à partir de ceux-ci, 34 points de repères ont été sélectionnés et 35 mesures ont été calculées. Les variables qui ne sont pas nécessaire (sexe, location géographique, sous-espèce), ont été contrôlées afin d’éviter des erreurs d’interprétation. Finalement, des matrices de corrélations et de variance/covariance (V/CV) ont été comparées dans chacun des ordres. Dans la seconde expérimentation, une étape s’ajoute, soit de comparer l’effet de la taille sur la présence de modules.
Plusieurs modules hypothétiques ont été testés. Dans la 1re hypothèse les cinq modules sont la base crânienne, la cavité crânienne, la région nasale, la région orale et la région zygomatique et forment les régions fonctionnelles du crâne. Dans la 2e hypothèse, le neurocrâne et le visage forment des modules distincts de développement majeur. La 3e hypothèse est que tous les traits neurocrâniens et faciaux sont séparés en deux modules distincts. Finalement, la 4e hypothèse, il existerait une matrice qui combinerait les 5 sous-régions du crâne en un seul module.
Résultats des études
Les matrices de corrélations et de V/CV entre les ordres ont démontré des niveaux modérés à élevés de similarité. Cela indique que les modules crâniens des mammifères sont restés relativement similaires au cours de leur diversification. Ce sont les changements dans l’intégration, c’est-à-dire dans le maintien de la forme, qui vont créer une différence entre les groupes. Dans la seconde étude, les chercheurs ont déterminé que si la croissance postnatale est plus élevée, comme chez les rongeurs et les marsupiaux, il y aura un plus grand pourcentage de variation de V/CV.
Les métathériens présentent une forte intégration des régions orale et nasale et presque aucune intégration au niveau du neurocrâne. L’hypothèse la plus plausible est que, puisque la survie du nouveau-né est en lien direct avec sa capacité à téter, son développement facial doit respecter une forme précise.
Contrairement au métathériens, la plupart des euthériens ont une intégration plus robuste au niveau du neurocrâne. Puisque leur période intra-utérine est plus longue, le système nerveux central a le temps de se développer avant que le besoin de téter soit présent. Pour ce qui est du genre Homo plus précisément, leur neurocrâne est bien intégré, mais le restant du crâne l’est moins. Cela pourrait indiquer que les humains ont été plus sensibles que les autres mammifères à la sélection naturelle. C’est ce qui ressort aussi de la deuxième étude ; les primates (et les chiroptères) ont un indice de flexibilité élevé au niveau de leurs schémas modulaires.
La tendance générale qui ressort de la phylogénie est une augmentation de la différenciation et une diminution de l’intégration des modules. Les espèces qui sont apparues plus récemment semblent s’adapter plus facilement à la sélection naturelle.
Conclusion
Les deux études en viennent à répondre aux trois questions initiales, soit s’il existe des modules similaires chez tous les mammifères, si ceux-ci sont en lien avec une fonction particulière et si la taille a une influence sur la détection des modules et leur flexibilité.
D’abord, il est possible de constater qu’il existe des modules assez similaires chez les mammifères. Ces schémas semblent englober les similitudes phénotypiques et génétiques sous-jacentes de chacune des espèces.
Ensuite, puisque le développement facial des métathériens et des euthériens est différents, il est possible de déduire qu’il existe un lien entre les modules et la fonction, dans ce cas-ci, l’alimentation à la naissance.
Finalement, la variation de la taille a pour effet de rendre les modules moins évidents à détecter et moins réceptif à la sélection naturelle. C’est le cas des marsupiaux, qui ont une croissance postnatale élevée. Au contraire, les humains qui ont une plus petite croissance postnatale, ont une plus grande flexibilité et donc une meilleure adaptation.
Modularité chez les insectes
Les insectes possèdent plusieurs segments, au niveau de leurs corps, indépendants les uns des autres. Ces segments peuvent avoir différentes fonctions et subir des modifications, sans influencer le reste de l'organisme. Ceci permet de mieux s'adapter aux pressions de l'environnement, en explorant divers arrangement spatial, hétérotopie, arrangement morphométrique des caractères, hétérométrie, ou arrangement du type de caractère pour un même segment, hétérotypie, niveau du corps de l'insecte pour mieux répondre aux pressions du milieu.
Ex. la quantité d'ailes présentes sur le corps ou le nombre d'appendices servant à la locomotion[16].
Organismes entiers holométaboles et hémimétaboles
Il est d’abord important de différencier les organismes hémimétaboles des holométaboles. Les organismes hémimétaboles sont ceux qui ont un développement direct, ne passent pas par un stade de larve avant d’atteindre le stade adulte. Ces organismes ont donc un stade juvénile où l’organisme est une copie très semblable à l’adulte, mais généralement en moins adapté et moins gros. Les organismes holométaboles sont ceux qui passent par des stades de développement qui passent par un stade larvaire avant de donner l’adulte. La physionomie du juvénile et de l’adulte sont particulièrement différentes l’une de l’autre[13]. Ces études ont permis d'en apprendre plus sur la modularité du développement, mais n'ont toutefois pas donné de réponse précise en ce qui concerne les variations du développement dû à la modularité. Il y a tellement de facteurs qui influencent le développement d'un organisme que ces changements sont difficilement attribuables, hors de tout doute, à une seule source[17].
Procédé de transcription
Le procédé de transcription est un bon exemple de modularité. Chaque site d’initiation recrute ses propres facteurs de transcription indépendamment des autres sites d’initiation. Cet exemple s’applique aussi pour les cascades de signalisation telles que Hedgehog, Wnt et Notch. Le réseau de régulation des gènes et les cascades de signalisation sont des exemples concrets de la modularité au niveau cellulaire. Si l’on pousse cet exemple plus loin, on obtient alors les interactions entre les différents tissus d’un organisme. La connexion avec les autres réseaux, dans d’autres tissus, s'effectue, entre autres, par les voies de signalisation ou cascade de signalisation, qui crée alors une autre réaction dans un autre endroit de l’organisme[7].
Il en est de même pour plusieurs récepteur qui utilisent les protéines modulaires lors de la signalisation entre les différentes cellules, mais il semblerait que certains récepteur ne nécessiteraient pas de protéine G pour la signalisation inter cellule.
Ces récepteurs couplés aux protéines G fonctionneraient alors selon une autre voie de transduction indépendante des protéines modulaires[18].
Éléments de démonstration
La théorie voulait que des réseaux construites en structure modulaire seraient plus résilients en cas de perturbation, ou éviterait le collapsus global. Ceci est longtemps resté non testé[3].
Une étude publiée en 2017 a testé les effets de la modularité sur la réponse aux perturbations au sein d'une métapopulation de microarthropodes construite en laboratoire et exposé à des perturabations[3].
Voir aussi
Articles connexes
Liens externes
- Bolker, J.A. (2000) Modularity in Development and Why It Matters to Evo-Devo. Integrative and Comparative biology, vol. 40, issue 5, pp. 770-776. https://doi.org/10.1093/icb/40.5.770
- Burgun, I. (2017) Évolution : une histoire de poissons. Agence Science Presse. https://www.sciencepresse.qc.ca/actualite/2017/10/11/evolution-histoire-poissons
Bibliographie
- Gilarranz L.J, Rayfield B, Liñán-Cembrano G, Bascompte J & Gonzalez A (2017), Effects of network modularity on the spread of perturbation impact in experimental metapopulations ; Science 14 Jul 2017:Vol. 357, Issue 6347, pp. 199-201 | DOI: 10.1126/science.aal4122
- Sales-Pardo M (2017) The importance of being modular ; Science 14 Jul 2017:Vol. 357, Issue 6347, pp. 128-129 DOI: 10.1126/science.aan8075
Références
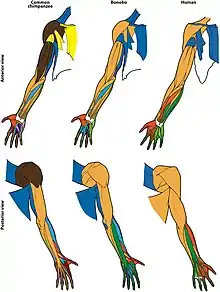
- (en) Rui Diogo, Julia L. Molnar, Campbell Rolian & Borja Esteve-Altava, « First anatomical network analysis of fore- and hindlimb musculoskeletal modularity in bonobos, common chimpanzees, and humans », Scientific Reports, vol. 8, no 6885, (DOI 10.1038/s41598-018-25262-6)
- Frelin, C., Swynghedauw, B. (2011). Biologie de l’évolution et médecine. Paris : Lavoisier. p.162, 178 pages.
- Gilarranz L.J, Rayfield B, Liñán-Cembrano G, Bascompte J & Gonzalez A (2017), Effects of network modularity on the spread of perturbation impact in experimental metapopulations ; Science 14 Jul 2017:Vol. 357, Issue 6347, pp. 199-201 | DOI: 10.1126/science.aal4122
- Schlosser, G., Wagner, G. P. (2004). Modularity in Development and Evolution. Chicago : University of Chicago Press. p.5-19, 600 pages.
- (en) Jessica A. Bolker, « Modularity in Development and Why It Matters to Evo-Devo », Integrative and Comparative Biology, vol. 40, no 5, , p. 770–776 (ISSN 1540-7063, DOI 10.1093/icb/40.5.770, lire en ligne, consulté le )
- Wagner G. P., (1996) Homologues, Natural Kinds and the Evolution of Modularity. American Zoologist, 36 (36-43), page38. Repéré à http://icb.oxfordjournals.org/content/36/1/36.short
- modularity in developpement and evolution.
- Caianiello S., (2011) Les modules de la variation : l’évo-dévo ou la nouvelle genèse des formes. Critique : Bodybuilding, l’évolution des corps, 1-2 (764-765), page13, 96 pages. Repéré à http://www.cairn.info/revue-critique-2011-1.htm#
- Amundson, R. (2005). The Changing Role of the Embryo in Evolutionnary Tought : Roots of Evo-Devo. New-York : Cambridge University Press. p. 244.
- Ambrose, B. A., Purugganan, M. D. (2012).Annual Plant Reviews, The Evolution of Plant Form. West Sussex : John Wiley & Sons. p. 280. 400 pages
- Sertier, A.-S. (2011). L’évolution modulaire des protéines : un point de vue phylogénétique (Mémoire de maitrise, Université de Lyon). Repéré à https://tel.archives-ouvertes.fr/tel-00842255
- Lozinguez-Giry, C., Kleman, J.P., Van der Rest, M. (1994). Modules et interactions moléculaires au sein des matrices extracellulaires. Médecine /Science, 10 (1234-43), page 1 – 3.
- Gilbert, S. F. (2004). Biologie du développement. Bruxelles : De Boeck Supérieur. p. 584, 767-768, 858 pages.
- (en) Arthur Porto, Felipe B. de Oliveira, Leila T. Shirai et Valderes De Conto, « The Evolution of Modularity in the Mammalian Skull I: Morphological Integration Patterns and Magnitudes », Evolutionary Biology, vol. 36, no 1, , p. 118–135 (ISSN 1934-2845, DOI 10.1007/s11692-008-9038-3, lire en ligne, consulté le )
- (en) Arthur Porto, Leila Teruko Shirai, Felipe Bandoni de Oliveira et Gabriel Marroig, « Size Variation, Growth Strategies, and the Evolution of Modularity in the Mammalian Skull », Evolution, vol. 67, no 11, , p. 3305–3322 (ISSN 1558-5646, DOI 10.1111/evo.12177, lire en ligne, consulté le )
- Raven, P. H., Johnson, G. B., Mason, K. A., Losos, J. B., Singer, S. S. (2011). Biologie. Bruxelles : De Boeck Supérieur. p. 679, 1406 pages.
- Maclaurin, J., Sterelny, K. (2008).What is Biodiversity?. Chicago: University of Chicago Press. p. 103, 224 pages.
- Bouvier M., Laporte S. A., Lagacé M., Caron M. G. (2000) Échafaudages protéiques et signalisation modulaire via les récepteurs à sept domaines transmembranaires : au-delà des protéines G. Médecine/sciences, 16 (644-51), page 645 & 650. Repéré à http://www.ipubli.inserm.fr/bitstream/handle/10608/2054/MS_2000_5_644.pdf?sequence=1