Lolitrème B
Le lolitrème B est un indole-diterpène trémorgène (qui cause des tremblements), considéré comme le principal responsable de la toxicité du ray-grass endophyté. C'est l'une des nombreuses mycotoxines produites par une espèce de champignons, Epichloë festucae var. lolii, qui parasite une graminées fourragère, le ray-grass anglais (Lolium perenne). Ce champignon endophyte vit en symbiose avec le ray-grass, il ne nuit pas à la plante, les toxines qu'il produit tuent les insectes qui se nourrissent aux dépens du ray-grass. Le lolitrème B est l'une de ces toxines, mais il est également nocif pour les mammifères. Les pousses et les fleurs du ray-grass infecté ont des concentrations particulièrement élevées de lolitrème B, et quand le bétail en ingère une trop grande quantité, il contracte la maladie dite « tétanie du ray-grass ». A faibles doses, les animaux ont des tremblements, et à des doses plus élevées ils chancellent, puis, à des doses encore plus élevées, les animaux souffrent de paralysie et meurent. Leur pression artérielle s'élève également. Les effets du lolitrème B apparaissent lentement et disparaissent de même, parce qu'il est stocké dans les graisses après ingestion.
| Lolitrème B | |
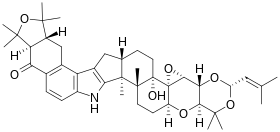
| |
| Identification | |
|---|---|
| Nom UICPA | (3S,4aR,4bR,5aS,5bS,16bS,18aS,19aS)-5b-hydroxy-
1,1,10,10,12,12,16b,16c-octaméthyl-3-(2-méthyl-1-propén -1-yl)-1,4a,4b,6,7,7a,9,9a,10,12,12a,16,16b,16c,17,18, 18a,19a-octadécahydro-5bH-[2]benzofuro[5,6-e][1,3]dioxino [5,4:2',3']oxiréno[4',4a']chroméno[5',6':6,7]indéno [1,2-b]indol-13(8H)-one |
| No CAS | |
| PubChem | 11947793 |
| SMILES | |
| InChI | |
| Propriétés chimiques | |
| Formule | C42H55NO7 |
| Masse molaire[1] | 685,888 6 ± 0,039 8 g/mol C 73,55 %, H 8,08 %, N 2,04 %, O 16,33 %, |
| Unités du SI et CNTP, sauf indication contraire. | |
Cette affection est particulièrement fréquente en Nouvelle-Zélande et en Australie, et les obtenteurs ont essayé de sélectionner des souches de champignons qui produisent des toxines plus sélectives, nuisibles aux ravageurs et non aux mammifères.
Le lolitrème B affecte une catégorie de canaux ioniques appelés canaux BK. Ces canaux s'ouvrent normalement temporairement pour permettre aux neurones et aux autres cellules électrosensibles, comme certaines cellules cardiaques, de se « réinitialiser » après qu'elles ont été excitées ; le lolitrème B les bloque, empêchant le neurone ou la cellule cardiaque de s'activer à nouveau. Cela affecte les fonctions nerveuses et cardiaques. Les canaux sont également impliqués dans le relâchement des vaisseaux sanguins, et le blocage des canaux provoque la constriction des vaisseaux sanguins, ce qui entraîne une augmentation de la pression artérielle.
Sources
Le lolitrème B se trouve dans les plants de ray-grass anglais (Lolium perenne) infectés par le champignon Epichloë festucae var. Lolii[2] - [3]. Ce champignon est un endophyte qui vit, pendant une partie de son cycle de vie, à l'intérieur des plantes, ses hyphes croissant entre les cellules végétales ; il est plus fréquent dans les tiges ou talles du ray-grass[2]. Le champignon produit du lolitrème B, l'une des nombreuses mycotoxines qui tuent les ravageurs, mais qui peut aussi être neurotoxique pour les mammifères[2].
Toxicité
Lorsque les animaux ingèrent des tiges de ray-grass infectées par Epichloë festucae var. Lolii, ils contractent une maladie appelée « tétanie du ray-grass. En cas d'intoxication légère, les animaux ont des tremblements, et, en cas d'intoxication plus sévère, ils chancellent et s'effondrent[2] - [4].
Chez les chevaux, on observe des tremblements des muscles du globe oculaire, qui sont plus sévères pendant les périodes d'alimentation et d'exercice[5]. Le lolitrème B peut également augmenter la fréquence cardiaque, la pression artérielle, le taux de respiration et perturber le processus de digestion[6] - [7]. Les lolitrèmes se distinguent parmi les neurotoxines trémorgènes parce qu'elles induisent un effet durable sur la fonction motrice et la fréquence cardiaque. Les tremblements peuvent durer des heures et à des concentrations élevées, ils peuvent causer la mort. Chez les animaux, les cas mortels dus au lolitrème B sont le plus souvent liés à des accidents malheureux tels que la chute dans un étang. Les effets neurotoxiques peuvent être complètement inversés[8].
Le seuil de toxicité varie d'une espèce à l'autre : chez les moutons, on a trouvé une valeur seuil de 1,8 à 2,0 mg/kg, et chez les bovins de 1,55 mg/kg[4] - [8]. La mesure de la concentration de lolitrème B dans le tissu adipeux peut être utilisée pour estimer la quantité de lolitrème B consommée et sert à déterminer la cause de la mort des bovins présentant des symptômes neurologiques[7] - [9]. Le lolitrème B agit probablement en synergie avec l'ergotamine pour augmenter la contraction des muscles lisses[10].
Épidémiologie
Epichloë festucae var. lolii infecte le ray-grass dans le monde entier, mais les cas de tétanie due au ray-grass sont rares en dehors de l'Australie et de la Nouvelle-Zélande. Les raisons de cet état de fait ne sont pas claires mais ont peut-être un rapport avec la sélection délibérée de ray-grass infectés par les endophytes par les obtenteurs, qui prennent en compte leur résistance aux ravageurs les plus répandus en Australie et en Nouvelle-Zélande, et avec la pratique de la monoculture par les agriculteurs de ces pays[2].
Prévention
Les sélectionneurs ont travaillé avec des mycologues en Australie et en Nouvelle-Zélande pour créer des souches de champignons produisant des mycotoxines toxiques pour les ravageurs mais pas pour les mammifères[2] En attendant que ces souches soient commercialisées, la meilleure prévention est d'éviter de faire paître le bétail sur du ray-grass lors de l'émergence des jeunes tiges et pendant la floraison (les concentrations les plus élevées se trouvent dans l'inflorescence épanouie et à la base des tiges), et en évitant le surpâturage. Dès que l'exposition au lolitrème B est terminée, les symptômes diminuent progressivement[2].
Pharmacologie
Le lolitrème B est rapidement éliminé du sérum et a une demi-vie de 14 minutes[11]. Le lolitrème B n'est pas très soluble et est généralement stocké dans la graisse après ingestion et libéré lentement. C'est probablement pourquoi ses effets se manifestent lentement et persistent après la fin de l'ingestion[2]. Plus la quantité ingérée est importante, plus il est stocké dans la graisse[12] - [13].
Le lolitrème B cible les canaux potassiques à grande conductivité activés par le calcium (canaux BK) et en particulier la sous-unité α (hSlo) des canaux BK[2]. Ces canaux sont ouverts temporairement pour permettre aux neurones de se « réinitialiser » après avoir été excités. Le lolitrème B les bloque, empêchant les neurones de s'exciter à nouveau après leur dépolarisation, ce qui entraîne des tremblements à faibles doses et, à doses élevées, la paralysie et la mort[2].
Le site de liaison du lolitrème B est susceptible d'être localisé dans cette sous-unité α[14] - [15]. Lorsque le lolitrème B est ajouté, le courant de potassium est rapidement supprimé et cette inhibition ne peut pas être inversée par lavage (cette inversion est possible pour la paxilline). Cependant, au fil du temps, le lolitrème B se dissocie lentement du site de liaison. L'inhibition par le lolitrème B dépend de la concentration en calcium. On a constaté que la concentration nécessaire pour une inhibition à 50 % de l'inhibition maximale (IC50) pour hSlo était de 3,7 ± 0,4 nM. Le lolitréme B est une neurotoxine plus puissante in vitro que la paxilline[11] - [14] - [15].
Le lolitrème B bloque de préférence la configuration ouverte des canaux BK, car sous des concentrations de calcium élevées favorisant l'ouverture des canaux BK, l'affinité apparente augmente d'un facteur trois[16] - [17]. L'inhibition par le lolitrème B et son affinité diffèrent en fonction de la concentration en calcium. Le lolitrème B a la plus grande affinité pour les canaux BK quand il y a une forte probabilité d'une conformation ouverte, par conséquent lorsque le calcium se lie aux sites à haute affinité. L'inhibition se produit lorsque les canaux sont à l'état ouvert[16].
Les canaux BK s'opposent à la vasoconstriction des vaisseaux sanguins entraînant une vasorelaxation. Le blocage des canaux conduit à une vasoconstriction et à une augmentation de la tension artérielle[18]. La sous-unité α du canal BK est exprimée dans les tissus musculaires et nerveux et les canaux BK sont abondants dans le cerveau[19]. Les canaux BK modulent la libération des neurotransmetteurs, la forme du potentiel d'action et les excitations répétitives[20]. L'inhibition des canaux peut expliquer pourquoi il se produit une libération accrue des neurotransmetteurs, entraînant des tremblements, une ataxie, une hypersensibilité, une augmentation de la contraction des muscles lisses du côlon et une fréquence cardiaque accrue[6] - [10] - [21]
Chimie
Le lolitrème B est le membre le plus puissant de la famille des lolitrèmes. Il possède une unité indole-diterpène ainsi qu'un groupe réactif époxyde[22].
Il ressemble structurellement à la paxilline, composé apparenté, également inducteur de tremblements[23]. Il existe plusieurs lolitrèmes qui sont désignés par une lettre différente. Ils diffèrent entre eux par la position et le nombre de substituants aryle et hydroxyle, ainsi que par l'absence ou la présence d'un cycle I. L'anneau I semble être nécessaire pour que des tremblements prolongés se produisent[24]. Les métabolites intermédiaires tels que les terpendoles et la paspaline peuvent devenir des lolitrèmes par addition de deux cycles (A et B) en position C20-C21 du groupe indole de la molécule[25].
Biosynthèse
La production de lolitrème B nécessite 10 gènes différents sur un seul locus (le locus LTM) qui est organisé en trois groupes. Ces groupes sont séparés par de longues séquences riches en AT. Le groupe 1 contient les gènes ltmG, ltmK et ltmM. Le groupe 2 les gènes ltmP, ltmF, ltmB, ltmQ et ltmC et le groupe 3 les gènes ltmE et ltmJ. Quatre gènes du groupe 2 sont des orthologues de gènes fonctionnels caractérisés de la paxilline, ce qui signifie que les gènes présentent des séquences homologues. Les gènes du groupe 3 semblent être spécifiques au genre Epichloë[26] - [27].
Notes et références
- Masse molaire calculée d’après « Atomic weights of the elements 2007 », sur www.chem.qmul.ac.uk.
- (en) G Philippe, « Lolitrem B and Indole Diterpene Alkaloids Produced by Endophytic Fungi of the Genus Epichloë and Their Toxic Effects in Livestock », Toxins, vol. 8, no 2, , p. 47 (DOI 10.3390/toxins8020047).
- (en) Adrian Leuchtmann, Charles W. Bacon, Christopher L. Schardl, James F. White et Mariusz Tadych, « Nomenclatural realignment of Neotyphodium species with genus Epichloë », Mycologia, vol. 106, no 2, , p. 202–215 (DOI 10.3852/13-251, lire en ligne).
- (en) P R Cheeke, « Endogenous toxins and mycotoxins in forage grasses and their effects on livestock », Journal of Animal Science, vol. 73, no 3, , p. 909 (ISSN 0021-8812, DOI 10.2527/1995.733909x, lire en ligne).
- (en) L. K. Johnstone, I. G. Mayhew et L. R. Fletcher, « Clinical expression of lolitrem B (perennial ryegrass) intoxication in horses », Equine Veterinary Journal, vol. 44, no 3, , p. 304–309 (ISSN 2042-3306, DOI 10.1111/j.2042-3306.2011.00439.x, lire en ligne).
- (en) L. M McLEAY, B. L SMITH et S. C MUNDAY-FINCH, « Tremorgenic mycotoxins paxilline, penitrem and lolitrem B, the non-tremorgenic 31-epilolitrem B and electromyographic activity of the reticulum and rumen of sheep », Research in Veterinary Science, vol. 66, no 2, , p. 119–127 (DOI 10.1053/rvsc.1998.0253, lire en ligne).
- (en) M. J. Fisher, D. W. Bohnert, C. J. Ackerman, C. S. Schauer, T. DelCurto, A. M. Craig et E. S. Vanzant, « Evaluation of perennial ryegrass straw as a forage source for ruminants », Journal of Animal Science, vol. 82, no 7, , p. 2175 (ISSN 1525-3163, DOI 10.2527/2004.8272175x, lire en ligne).
- (en) R. T. Gallagher et A. D. Hawkes, « The potent tremorgenic neurotoxins lolitrem B and aflatrem: A comparison of the tremor response in mice », Experientia, vol. 42, no 7, , p. 823–825 (ISSN 0014-4754, DOI 10.1007/BF01941539, lire en ligne).
- (en) J. Tor-Agbidye, L. L. Blythe et A. M. Craig, « Correlation of endophyte toxins (ergovaline and lolitrem B) with clinical disease: fescue foot and perennial ryegrass staggers », Veterinary and Human Toxicology, vol. 43, no 3, , p. 140–146 (ISSN 0145-6296).
- (en) J. E. Dalziel, K. E. Dunstan et S. C. Finch, « Combined effects of fungal alkaloids on intestinal motility in an in vitro rat model », Journal of Animal Science, vol. 91, no 11, , p. 5177–5182 (ISSN 1525-3163, DOI 10.2527/jas.2013-6449, lire en ligne).
- D. Grancher, A. Durix, Y. Moulard, Y. Bonnaire, M. Caecelen, Y. Camier et S. Bony, « ''Distribution du lolitrème B et de l’ergovaline après un bolus intraruminal chez la chèvre en lactation », Rencontres Recherches Ruminants, vol. 11, (lire en ligne).
- (en) Shigeru Miyazaki, Ikumi Ishizaki, Masumi Ishizaka, Tadashi Kanbara et Yuriko Ishiguro-Takeda, « Lolitrem B Residue in Fat Tissues of Cattle Consuming Endophyte-Infected Perennial Ryegrass Straw », Journal of Veterinary Diagnostic Investigation, vol. 16, no 4, , p. 340–342 (ISSN 1040-6387, DOI 10.1177/104063870401600416, lire en ligne).
- (en) S. C. Finch, L. R. Fletcher et J. V. Babu, « The evaluation of endophyte toxin residues in sheep fat », New Zealand Veterinary Journal, vol. 60, no 1, , p. 56–60 (ISSN 0048-0169, DOI 10.1080/00480169.2011.634746, lire en ligne).
- (en) Julie E. Dalziel, Sarah C. Finch et James Dunlop, « The fungal neurotoxin lolitrem B inhibits the function of human large conductance calcium-activated potassium channels », Toxicology Letters, vol. 155, no 3, , p. 421–426 (DOI 10.1016/j.toxlet.2004.11.011, lire en ligne).
- (en) Wendy L. Imlach, Sarah C. Finch, James Dunlop, Andrea L. Meredith, Richard W. Aldrich et Julie E. Dalziel, « The Molecular Mechanism of "Ryegrass Staggers," a Neurological Disorder of K Channels », Journal of Pharmacology and Experimental Therapeutics, vol. 327, no 3, , p. 657–664 (DOI 10.1124/jpet.108.143933, lire en ligne).
- (en) Wendy L. Imlach, Sarah C. Finch, Yanli Zhang, James Dunlop et Julie E. Dalziel, « Mechanism of action of lolitrem B, a fungal endophyte derived toxin that inhibits BK large conductance Ca2+-activated K+ channels », Toxicon, vol. 57, no 5, , p. 686–694 (DOI 10.1016/j.toxicon.2011.01.013, lire en ligne).
- (en) Xu-Hui Zeng, Xiao-Ming Xia et Christopher J. Lingle, « Divalent Cation Sensitivity of BK Channel Activation Supports the Existence of Three Distinct Binding Sites », The Journal of General Physiology, vol. 125, no 3, , p. 273–286 (ISSN 0022-1295, DOI 10.1085/jgp.200409239, lire en ligne).
- (en) J. E. Brayden et M. T. Nelson, « Regulation of arterial tone by activation of calcium-dependent potassium channels », Science, vol. 256, no 5056, , p. 532–535 (ISSN 0036-8075, DOI 10.1126/science.1373909, lire en ligne).
- (en) H.G. Knaus, « Distribution of high-conductance Ca2+-activated K+ channels in rat brain: targeting to axons and nerve terminals », The Journal.of Neuroscience, vol. 16, (lire en ligne).
- (en) Li-Rong Shao, Ragnhild Halvorsrud, Lyle Borg-Graham et Johan F. Storm, « The role of BK-type Ca2+-dependent K+ channels in spike broadening during repetitive firing in rat hippocampal pyramidal cells », The Journal of Physiology, vol. 521, no 1, , p. 135–146 (ISSN 1469-7793, DOI 10.1111/j.1469-7793.1999.00135.x, lire en ligne).
- (en) P.G. Mantle, « Amino acid neurotransmitter release from cerebrocortical synaptosomes of sheep with severe ryegrass staggers in New Zealand », Research in Veterinary Science, vol. 34, .
- (en) R. T. Gallagher, E. P. White et P. H. Mortimer, « Ryegrass Staggers: Isolation of Potent Neurotoxins Lolitrem a and Lolitrem B From Staggers-Producing Pastures », New Zealand Veterinary Journal, vol. 29, no 10, , p. 189–190 (ISSN 0048-0169, DOI 10.1080/00480169.1981.34843, lire en ligne).
- (en) Christopher O. Miles, Alistair L. Wilkins, Rex T. Gallagher, Allan D. Hawkes, Sarah C. Munday et Neale R. Towers, « Synthesis and tremorgenicity of paxitriols and lolitriol: possible biosynthetic precursors of lolitrem B », Journal of Agricultural and Food Chemistry, vol. 40, no 2, , p. 234–238 (ISSN 0021-8561, DOI 10.1021/jf00014a013, lire en ligne).
- (en) Sarah C. Munday-Finch, Alistair L. Wilkins, Christopher O. Miles, Hiroshi Tomoda et Satoshi Ōmura, « Isolation and Structure Elucidation of Lolilline, a Possible Biosynthetic Precursor of the Lolitrem Family of Tremorgenic Mycotoxins », Journal of Agricultural and Food Chemistry, vol. 45, no 1, , p. 199–204 (ISSN 0021-8561, DOI 10.1021/jf960396r, lire en ligne).
- (en) Carolyn A. Young, Brian A. Tapper, Kimberley May, Christina D. Moon, Christopher L. Schardl et Barry Scott, « Indole-Diterpene Biosynthetic Capability of Epichloë Endophytes as Predicted by ltm Gene Analysis », Applied and Environmental Microbiology, vol. 75, no 7, , p. 2200–2211 (ISSN 0099-2240, DOI 10.1128/AEM.00953-08, lire en ligne).
- (en) Carolyn A. Young, Silvina Felitti, Katherine Shields, German Spangenberg, Richard D. Johnson, Gregory T. Bryan et Sanjay Saikia, « A complex gene cluster for indole-diterpene biosynthesis in the grass endophyte Neotyphodium lolii », Fungal Genetics and Biology, vol. 43, no 10, , p. 679–693 (DOI 10.1016/j.fgb.2006.04.004, lire en ligne).
- (en) C. A. Young, M. K. Bryant, M. J. Christensen, B. A. Tapper, G. T. Bryan et B. Scott, « Molecular cloning and genetic analysis of a symbiosis-expressed gene cluster for lolitrem biosynthesis from a mutualistic endophyte of perennial ryegrass », Molecular Genetics and Genomics, vol. 274, no 1, , p. 13–29 (ISSN 1617-4615, DOI 10.1007/s00438-005-1130-0, lire en ligne).
Voir aussi
Articles connexes
- Tournis du ray-grass
- Défense des plantes par champignons endophytes
- Pénitrème A, neurotoxine fongique structurellement apparentée au lolitrème B, également présente chez le ray-grass