Domestication des animaux
La domestication des animaux est la relation mutuelle entre les animaux non humains et les humains qui ont une influence sur leurs soins et leur reproduction[1].

Charles Darwin a reconnu un petit nombre de traits qui rendaient les espèces domestiquées différentes de leurs ancêtres sauvages. Il a également été le premier à reconnaître la différence entre l'élevage sélectif conscient (c'est-à-dire la sélection artificielle) dans lequel les humains sélectionnent directement les traits désirables, et la sélection inconsciente où les traits évoluent comme un sous-produit de la sélection naturelle ou de la sélection sur d'autres traits[2] - [3] - [4]. Il existe une différence génétique entre les populations domestiques et sauvages. Il existe également une différence génétique entre les traits de domestication qui, selon les chercheurs, ont été essentiels aux premiers stades de la domestication, et les traits d'amélioration apparus depuis la scission entre les populations sauvages et domestiques[5] - [6] - [7]. Les traits de domestication sont généralement fixés dans tous les animaux domestiques et ont été sélectionnés lors de l'épisode initial de domestication de cet animal ou de cette plante, alors que les traits d'amélioration ne sont présents que dans une partie des animaux domestiques, bien qu'ils puissent être fixés dans des races individuelles ou des populations régionales[6] - [7] - [8].
La domestication ne doit pas être confondue avec l'apprivoisement. L'apprivoisement est la modification comportementale conditionnée d'un animal né dans la nature lorsque son évitement naturel des humains est réduit et qu'il accepte la présence des humains, mais la domestication est la modification génétique permanente d'une lignée élevée qui conduit à une prédisposition héréditaire envers les humains[9] - [10] - [3]. Certaines espèces animales, et certains individus au sein de ces espèces, sont de meilleurs candidats à la domestication que d'autres car ils présentent certaines caractéristiques comportementales :
- la taille et l'organisation de leur structure sociale ;
- la disponibilité et le degré de sélectivité dans leur choix de partenaires ;
- la facilité et la rapidité avec lesquelles les parents se lient à leurs petits, et la maturité et la mobilité des petits à la naissance ;
- le degré de flexibilité dans le régime alimentaire et la tolérance à l'habitat ;
- les réponses aux humains et aux nouveaux environnements, y compris les réponses de fuite et la réactivité aux stimuli externes[11] - [12] - [9].
Il est proposé qu'il y ait trois voies principales que la plupart des animaux domestiques ont suivies dans la domestication :
- les commensaux, adaptés à une niche humaine (par exemple, les chiens, les chats, la volaille, éventuellement les porcs) ;
- les animaux recherchés pour la nourriture et autres sous-produits (par exemple, moutons, chèvres, bovins, buffles d'eau, yack, porc, renne, lama, alpaga et dinde) ;
- les animaux ciblés pour les ressources de trait et non alimentaires (par exemple, cheval, âne, chameau)[7] - [11] - [13] - [14] - [15].
Le chien a été le premier animal à être domestiqué[16] - [17] et s'est établi dans toute l'Eurasie avant la fin du Pléistocène supérieur, bien avant la culture et avant la domestication d'autres animaux[16]. Contrairement à d'autres espèces domestiques qui ont été principalement sélectionnées pour des traits liés à la production, les chiens ont d'abord été sélectionnés pour leurs comportements[18]. Les données archéologiques et génétiques suggèrent que le flux génétique bidirectionnel à long terme entre les stocks sauvages et domestiques -y compris les ânes, les chevaux, les camélidés du Nouveau et de l'Ancien Monde, les chèvres, les moutons et les porcs- était courant[7] - [14]. Une étude a conclu que la sélection humaine pour les traits domestiques a probablement contrecarré l'effet d'homogénéisation du flux génétique des sangliers vers les porcs et créé des « îlots de domestication » dans le génome. Le même processus peut également s'appliquer à d'autres animaux domestiques. Parmi des animaux les plus couramment domestiqués sont les chats et les chiens[19] - [20].
Histoire
La domestication des animaux et des plantes a été déclenchée par les changements climatiques et environnementaux qui se sont produits après le pic du dernier maximum glaciaire il y a environ 21 000 ans et qui se poursuivent jusqu'à nos jours. Ces changements ont rendu l'obtention de nourriture difficile. Le premier domestique était le chien domestique (Canis lupus familiaris) d'un ancêtre du loup (Canis lupus) il y a au moins 15 000 ans. Le Dryas jeune qui s'est produit il y a 12 900 ans était une période de froid intense et d'aridité qui a poussé les humains à intensifier leurs stratégies de recherche de nourriture. Au début de l'Holocène, il y a 11 700 ans, des conditions climatiques favorables et l'augmentation des populations humaines ont conduit à la domestication d'animaux et de plantes à petite échelle, ce qui a permis aux humains d'augmenter la nourriture qu'ils obtenaient grâce à la chasse-cueillette[21].
L'utilisation accrue de l'agriculture et la domestication continue des espèces au cours de la transition néolithique ont marqué le début d'un changement rapide dans l'évolution, l'écologie et la démographie des humains et de nombreuses espèces d'animaux et de plantes[22] - [7]. Les zones d'agriculture croissante ont subi une urbanisation[23], développé des populations à plus forte densité, développé des économies et sont devenues des centres de domestication du bétail et des cultures[24] - [25] - [26]. De telles sociétés agricoles ont émergé à travers l'Eurasie, l'Afrique du Nord et l'Amérique du Sud et centrale.
Dans le Croissant Fertile, il y a entre 10 000 à 11 000 ans, la zooarchéologie indique que les chèvres, les porcs, les moutons et les bovins taurins ont été les premiers animaux à être domestiqués. Des archéologues travaillant à Chypre ont trouvé un lieu d'enterrement plus ancien, vieux d'environ 9 500 ans, d'un humain adulte avec un squelette de félin. Deux mille ans plus tard, des zébus à bosse ont été domestiqués dans ce qui est aujourd'hui le Baloutchistan au Pakistan. En Asie de l'Est, il y a 8 000 ans, des porcs ont été domestiqués à partir de sangliers génétiquement différents de ceux trouvés dans le Croissant fertile. Le cheval a été domestiqué dans la steppe d'Asie centrale il y a 5 500 ans. Le poulet d'Asie du Sud-Est a été domestiqué il y a 4 000 ans[21].
Caractéristiques universelles
La biomasse des vertébrés sauvages est désormais de plus en plus faible par rapport à la biomasse des animaux domestiques, la biomasse calculée des seuls bovins domestiques étant supérieure à celle de tous les mammifères sauvages[27]. Parce que l'évolution des animaux domestiques est continue, le processus de domestication a un début mais pas une fin. Divers critères ont été établis pour fournir une définition des animaux domestiques, mais toutes les décisions concernant le moment exact où un animal peut être étiqueté « domestiqué » au sens zoologique sont arbitraires, bien que potentiellement utiles[28]. La domestication est un processus fluide et non linéaire qui peut démarrer, s'arrêter, s'inverser ou emprunter des chemins inattendus sans seuil clair ou universel séparant le sauvage du domestique. Cependant, il existe des caractéristiques universelles communes à tous les animaux domestiques[11].
Préadaptation comportementale
Certaines espèces animales, et certains individus au sein de ces espèces, sont de meilleurs candidats à la domestication que d'autres car ils présentent certaines caractéristiques comportementales :
- la taille et l'organisation de leur structure sociale ;
- la disponibilité et le degré de sélectivité dans leur choix de partenaires ;
- la facilité et la rapidité avec lesquelles les parents se lient à leurs petits, et la maturité et la mobilité des petits à la naissance ;
- le degré de flexibilité dans le régime alimentaire et la tolérance à l'habitat ;
- les réponses aux humains et aux nouveaux environnements, y compris les réponses de fuite et la réactivité aux stimuli externes[11] - [12] - [9].
Ces caractéristiques sont également la cible principale des pressions de sélection subies par l'animal en cours de domestication. Cela implique que tous les animaux ne peuvent pas être domestiqués, par ex. un membre sauvage de la famille des chevaux, le zèbre[7] - [29].
Jared Diamond, dans son livre Guns, Germs, and Steel, se demande pourquoi seuls 14 grands mammifères herbivores terrestres ont été domestiqués sur les 148 espèces présentes dans le monde. Il suggère que leurs ancêtres sauvages ont présenté les six caractéristiques suivantes pour leur domestication[3]:268-174 :
- un régime alimentaire efficace - ainsi les animaux herbivores sont les plus faciles à garder en captivité. Les carnivores, quant à eux, obligent à maintenir des animaux s'alimentant de plantes, pour les nourrir ce qui, au final, accroit la quantité totale de végétaux nécessaire ;
- un taux de croissance rapide - et ce de manière à ce que l'animal domestiqué soit le plus rapidement utile. Certains grands animaux nécessitent de grandes durées avant d'atteindre la taille utile ;
- une capacité à se reproduire en captivité - seuls les animaux pouvant se reproduire en captivité peuvent être domestiqués, les autres nécessiteront toujours des prélèvements dans la nature ;
- des disposition agréables - seuls les animaux présentant de bonnes dispositions envers l'humain peuvent être domestiqués, les autres présenteront des dangers potentiels ;
- une tendance à ne pas paniquer - certaines espèces sont nerveuses, rapides et sujettes à la fuite lorsqu'elles perçoivent une menace, elles ne seront pas propices à la domestication ;
- une structure sociale - toutes les espèces de grands mammifères domestiqués ont des ancêtres sauvages vivant en troupeaux régis par une hiérarchie de dominance. Ces troupeaux avaient des territoires d'origine qui se chevauchaient plutôt que des territoires d'origine mutuellement exclusifs. Cet arrangement a permis aux humains de prendre le contrôle de la hiérarchie de dominance.
Taille et fonction du cerveau
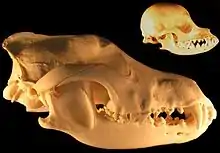
La sélection soutenue pour une réactivité réduite chez les mammifères domestiques a entraîné de profonds changements dans la forme et la fonction du cerveau. Plus la taille du cerveau est grande au départ et plus son degré de pliage est élevé, plus le degré de réduction de la taille du cerveau sous domestication est important[11] - [30]. Des renards élevés sélectivement pour la docilité pendant 40 ans ont connu une réduction significative de la hauteur et de la largeur crâniennes et par inférence de la taille de leur cerveau[11] - [31], ce qui étaye l'hypothèse selon laquelle la réduction de la taille du cerveau est une réponse précoce à la pression sélective pour la docilité et la réactivité réduite qui est une caractéristique universelle de la domestication des animaux. La partie du cerveau la plus touchée chez les mammifères domestiques est le système limbique qui, chez les chiens, les porcs et les moutons domestiques, présente une réduction de taille de 40 % par rapport à leurs espèces sauvages. Cette partie du cerveau régule la fonction endocrinienne qui influence les comportements tels que l'agressivité, la méfiance et les réponses au stress induit par l'environnement, tous des attributs qui sont considérablement affectés par la domestication[11] - [30].
Pléiotropie
Une cause putative des changements importants observés dans le syndrome de domestication est la pléiotropie. La pléiotropie se produit lorsqu'un gène influence deux ou plusieurs traits phénotypiques apparemment sans rapport. Certains changements physiologiques caractérisent les animaux domestiques de nombreuses espèces. Ces changements comprennent de nombreuses marques blanches (en particulier sur la tête), des oreilles tombantes et des queues bouclées. Celles-ci surviennent même lorsque la docilité est le seul trait sous pression sélective[32]. Les gènes impliqués dans l'apprivoisement sont largement inconnus, on ne sait donc pas comment ni dans quelle mesure la pléiotropie contribue au syndrome de domestication. L'apprivoisement peut être causé par la régulation à la baisse des réactions de peur et de stress via la réduction des glandes surrénales[32]. Sur cette base, les hypothèses de pléiotropie peuvent être séparées en deux théories. L'hypothèse de la crête neurale relie la fonction de la glande surrénale aux déficits des cellules de la crête neurale au cours du développement. L'hypothèse du réseau de régulation génétique unique affirme que les changements génétiques dans les régulateurs en amont affectent les systèmes en aval[33] - [34].
Les cellules de la crête neurale (NCC) sont des cellules souches embryonnaires de vertébrés qui fonctionnent directement et indirectement au début de l'embryogenèse pour produire de nombreux types de tissus. Étant donné que les traits couramment affectés par le syndrome de domestication sont tous dérivés du NCC en développement, l'hypothèse de la crête neurale suggère que les déficits de ces cellules provoquent le domaine des phénotypes observés dans le syndrome de domestication[34]. Ces déficits pourraient provoquer des changements chez de nombreux mammifères domestiques, tels que les oreilles coupées (observées chez le lapin, le chien, le renard, le porc, le mouton, la chèvre, le bétail et les ânes) ainsi que les queues bouclées (porcs, renards et chiens). Bien qu'elles n'affectent pas directement le développement du cortex surrénalien, les cellules de la crête neurale peuvent être impliquées dans des interactions embryologiques pertinentes en amont[33]. De plus, la sélection artificielle ciblant l'apprivoisement peut affecter les gènes qui contrôlent la concentration ou le mouvement des NCC dans l'embryon, conduisant à une variété de phénotypes[34].
L'hypothèse du réseau de régulation génétique unique propose que le syndrome de domestication résulte de mutations dans les gènes qui régulent le modèle d'expression de gènes plus en aval[32]. Par exemple, la coloration pie ou tachetée du pelage peut être causée par une liaison dans les voies biochimiques des mélanines impliquées dans la coloration du pelage et des neurotransmetteurs tels que la dopamine qui aident à façonner le comportement et la cognition[11] - [35]. Ces traits liés peuvent provenir de mutations dans quelques gènes régulateurs clés. Un problème avec cette hypothèse est qu'elle propose qu'il existe des mutations dans les réseaux de gènes qui provoquent des effets dramatiques qui ne sont pas mortels, mais aucun réseau de régulation génétique actuellement connu ne provoque un changement aussi spectaculaire dans tant de traits différents[33].
Réversion limitée
Les mammifères sauvages tels que les chiens, les chats, les chèvres, les ânes, les cochons et les furets qui ont vécu séparés des humains pendant des générations ne montrent aucun signe de récupération de la masse cérébrale de leurs progéniteurs sauvages[11] - [36]. Les dingos vivent séparés des humains depuis des milliers d'années mais ont toujours la même taille de cerveau que celle d'un chien domestique[11] - [37]. Les chiens sauvages qui évitent activement le contact humain dépendent toujours des déchets humains pour leur survie et ne sont pas revenus aux comportements autonomes de leurs ancêtres loups[11] - [38].
Voies de domestication
Depuis 2012, un modèle multi-étapes de domestication animale a été accepté par deux groupes. Le premier groupe a proposé que la domestication des animaux se déroule selon un continuum d'étapes allant de l'anthropophilie, du commensalisme, du contrôle dans la nature, du contrôle des animaux en captivité, de l'élevage extensif, de l'élevage intensif et enfin des animaux de compagnie dans une relation lente et s'intensifiant progressivement entre les humains et les animaux[39] - [40].

Le deuxième groupe a proposé qu'il y avait trois voies principales que la plupart des animaux domestiques ont suivies dans la domestication :
- les commensaux, adaptés à une niche humaine (par exemple, les chiens, les chats, la volaille, éventuellement les porcs) ;
- les animaux recherchés pour la nourriture et autres sous-produits (par exemple, moutons, chèvres, bovins, buffles d'eau, yack, porc, renne, lama, alpaga et dinde) ;
- les animaux ciblés pour les ressources de trait et non alimentaires (par exemple, cheval, âne, chameau)[7] - [11] - [13] - [14] - [15].
La voie commensale
La voie commensale était parcourue par des vertébrés qui se nourrissaient de déchets autour des habitats humains ou par des animaux qui se nourrissaient d'autres animaux attirés vers les camps humains. Ces animaux ont établi une relation commensale avec les humains dans laquelle les animaux ont bénéficié mais les humains n'ont reçu aucun mal mais peu d'avantages. Les animaux les plus capables de tirer parti des ressources associées aux camps humains auraient été les individus les plus dompteurs et les moins agressifs avec des distances de combat ou de fuite plus courtes[41] - [42] - [43]. Plus tard, ces animaux ont développé des liens sociaux ou économiques plus étroits avec les humains qui ont conduit à une relation domestique[7] - [11] - [19]. Le saut d'une population anthropophilique à une population domestique n'aurait pu avoir lieu qu'après que les animaux aient progressé de l'anthropophilie à l'accoutumance, au commensalisme et au partenariat, lorsque la relation entre l'animal et l'humain aurait jeté les bases de la domestication, y compris la captivité et la reproduction contrôlée par les hommes. De ce point de vue, la domestication animale est un processus coévolutif dans lequel une population répond à une pression sélective tout en s'adaptant à une nouvelle niche qui comprenait une autre espèce aux comportements évolutifs. Les animaux de la voie commensale comprennent les chiens, les chats, les volailles et éventuellement les porcs.
La domestication des animaux a commencé plus de 15 000 ans avant le présent (YBP), en commençant par le loup gris (Canis lupus) par des chasseurs-cueilleurs nomades. Ce n'est qu'en 11 000 YBP que les habitants du Proche-Orient ont noué des relations avec des populations sauvages d'aurochs, de sangliers, de moutons et de chèvres. Un processus de domestication a alors commencé à se développer. Le loup gris a très probablement suivi la voie commensale de la domestication. Quand, où et combien de fois les loups ont pu être domestiqués restent débattus car seul un petit nombre de spécimens anciens ont été trouvés, et l'archéologie et la génétique continuent de fournir des preuves contradictoires. Les restes de chiens les plus largement acceptés et les plus anciens remontent à 15 000 ans pour le chien de Bonn-Oberkassel. Des vestiges antérieurs datant de 30 000 ans ont été décrits comme des chiens paléolithiques, mais leur statut de chien ou de loup reste débattu. Des études récentes indiquent qu'une divergence génétique s'est produite entre les chiens et les loups entre 20 000 et 40 000 ans, mais il s'agit du délai supérieur de domestication car il représente le moment de la divergence et non le moment de la domestication[44].
Le poulet est l'une des espèces domestiquées les plus répandues et l'une des plus grandes sources de protéines au monde. Bien que le poulet ait été domestiqué en Asie du Sud-Est, des preuves archéologiques suggèrent qu'il n'a pas été élevé comme espèce d'élevage avant l'an 400 au Levant. Avant cela, les poulets étaient associés aux humains depuis des milliers d'années et gardés pour les combats de coqs, les rituels et les zoos royaux, ils n'étaient donc pas à l'origine une espèce de proie. Le poulet n'était pas un aliment populaire en Europe jusqu'à il y a seulement mille ans[45].
La voie des proies

La voie des proies était la voie par laquelle la plupart des principales espèces de bétail sont entrées dans la domestication, car elles étaient autrefois chassées par les humains pour leur viande. La domestication a probablement commencé lorsque les humains ont commencé à expérimenter des stratégies de chasse conçues pour augmenter la disponibilité de ces proies, peut-être en réponse à une pression localisée sur l'approvisionnement de l'animal. Au fil du temps et avec les espèces les plus réactives, ces stratégies de gestion du gibier se sont transformées en stratégies de gestion du troupeau qui comprenaient le contrôle multigénérationnel soutenu sur les mouvements, l'alimentation et la reproduction des animaux. Au fur et à mesure que l'interférence humaine dans les cycles de vie des proies s'intensifiait, les pressions évolutives pour un manque d'agressivité auraient conduit à l'acquisition des mêmes traits de syndrome de domestication trouvés chez les animaux domestiques commensaux[7] - [11] - [19].
Les animaux de la voie des proies comprennent les moutons, les chèvres, les bovins, les buffles d'eau, les yacks, les porcs, les rennes, les lamas et les alpagas. Les bonnes conditions pour la domestication de certains d'entre eux semblent avoir été en place dans le centre et l'est du Croissant fertile à la fin du ralentissement climatique du Dryas jeune et au début de l'Holocène précoce il y a environ 11 700 ans. Il y a 10 000 ans les gens tuaient préférentiellement les jeunes mâles d'une espèce donnée, ce qui permettait aux femelles de produire plus de progéniture. En mesurant la taille, les sex-ratios et les profils de mortalité des spécimens zooarchéologiques, les archéologues ont pu documenter les changements dans les stratégies de gestion des moutons, chèvres, porcs et vaches chassés dans le Croissant fertile à partir de 11 700 YBP. Une récente étude démographique et métrique des restes de vaches et de porcs à Sha'ar Hagolan, en Israël, a démontré que les deux espèces étaient sévèrement chassées avant la domestication, suggérant que l'exploitation intensive a conduit à des stratégies de gestion adoptées dans toute la région qui ont finalement conduit à la domestication de ces espèces. populations suivant la voie des proies. Ce schéma de chasse excessive avant la domestication suggère que la voie des proies était aussi accidentelle et non intentionnelle que la voie commensale[7] - [13].
La voie dirigée

La voie dirigée était un processus plus délibéré et dirigé initié par les humains dans le but de domestiquer un animal vivant en liberté. Il n'a probablement vu le jour qu'une fois que les gens se sont familiarisés avec les animaux domestiques commensaux ou proies. Ces animaux ne possédaient probablement pas bon nombre des préadaptations comportementales que certaines espèces présentent avant la domestication. Par conséquent, la domestication de ces animaux nécessite des efforts plus délibérés de la part des humains pour contourner les comportements qui ne facilitent pas la domestication, avec une assistance technologique accrue nécessaire[7] - [11] - [13].
Les humains dépendaient déjà des plantes et des animaux domestiques lorsqu'ils ont imaginé les versions domestiques des animaux sauvages. Bien que les chevaux, les ânes et les chameaux de l'Ancien Monde aient parfois été chassés comme espèces de proies, ils ont chacun été délibérément introduits dans la niche humaine pour les sources de transport. La domestication était encore une adaptation multigénérationnelle aux pressions de sélection humaines, y compris la docilité, mais sans une réponse évolutive appropriée, la domestication n'a pas été réalisée[7]. Par exemple, malgré le fait que les chasseurs de gazelle du Proche-Orient à l'Épipaléolithique évitaient d'abattre les femelles reproductrices pour favoriser l'équilibre de la population, ni les gazelles[7] - [26] ni les zèbres[7] - [46] ne possédaient les conditions préalables nécessaires et n'ont jamais été domestiqués. Il n'y a aucune preuve claire de la domestication d'un animal de proie en troupeau en Afrique, à l'exception notable de l'âne, qui a été domestiqué en Afrique du Nord-Est au cours du 4e millénaire avant notre ère[47].
Voies multiples
Les voies que les animaux ont pu suivre ne sont pas mutuellement exclusives. Les porcs, par exemple, peuvent avoir été domestiqués lorsque leurs populations se sont habituées à la niche humaine, ce qui suggérerait une voie commensale, ou ils peuvent avoir été chassés et suivre une voie de proie, ou les deux[7] - [11] - [13].
Flux de gènes post-domestication
Au fur et à mesure que les sociétés agricoles émigraient des centres de domestication en emmenant avec elles leurs partenaires domestiques, elles rencontraient des populations d'animaux sauvages de la même espèce ou d'espèces sœurs. Parce que les domestiques partageaient souvent un ancêtre commun récent avec les populations sauvages, ils étaient capables de produire une progéniture fertile. Les populations domestiques étaient petites par rapport aux populations sauvages environnantes, et des hybridations répétées entre les deux ont finalement conduit la population domestique à devenir génétiquement plus divergente de sa population source domestique d'origine[28] - [48].
Les progrès de la technologie de séquençage de l'ADN permettent d'accéder au génome nucléaire et de l'analyser dans un cadre de génétique des populations. La résolution accrue des séquences nucléaires a démontré que le flux de gènes est courant, non seulement entre des populations domestiques géographiquement diverses d'une même espèce, mais également entre des populations domestiques et des espèces sauvages qui n'ont jamais donné naissance à une population domestique[7].
- Le trait de patte jaune possédé par de nombreuses races de poulets commerciales modernes a été acquis par introgression de la sauvagine grise indigène d'Asie du Sud[7] - [49].
- Les bovins africains sont des hybrides qui possèdent à la fois un signal mitochondrial maternel des bovins taurins européens et une signature du chromosome Y paternel des bovins indiens asiatiques[7] - [50].
- De nombreuses autres espèces de bovidés, dont le bison, le yack, le banteng et le gaur, s'hybrident également facilement[7] - [51].
- Il a été démontré que les chats et les chevaux[52] s'hybrident avec de nombreuses espèces étroitement apparentées[7].
- Les abeilles domestiques se sont accouplées avec tant d'espèces différentes qu'elles possèdent maintenant des génomes plus variables que leurs ancêtres sauvages d'origine[7] - [53].
Les données archéologiques et génétiques suggèrent que le flux génétique bidirectionnel à long terme entre les stocks sauvages et domestiques - y compris les canidés, les ânes, les chevaux, les camélidés du Nouveau et de l'Ancien Monde, les chèvres, les moutons et les porcs - était courant. Le flux de gènes bidirectionnel entre les rennes domestiques et sauvages se poursuit aujourd'hui[7] - [14].
Les données archéologiques et génétiques suggèrent que le flux génétique bidirectionnel à long terme entre les stocks sauvages et domestiques - y compris les canidés, les ânes, les chevaux, les camélidés du Nouveau et de l'Ancien Monde, les chèvres, les moutons et les porcs - était courant. Le flux de gènes bidirectionnel entre les rennes domestiques et sauvages se poursuit aujourd'hui.
La conséquence de cette introgression est que les populations domestiques modernes peuvent souvent sembler avoir une affinité génomique beaucoup plus grande avec les populations sauvages qui n'ont jamais été impliquées dans le processus de domestication d'origine. Par conséquent, il est proposé que le terme "domestication" soit réservé uniquement au processus initial de domestication d'une population discrète dans le temps et dans l'espace. Le mélange ultérieur entre des populations domestiques introduites et des populations sauvages locales qui n'ont jamais été domestiquées doit être qualifié de "capture introgressive". Confondre ces deux processus brouille notre compréhension du processus original et peut conduire à une inflation artificielle du nombre de fois où la domestication a eu lieu[7] - [28]. Cette introgression peut, dans certains cas, être considérée comme une introgression adaptative, comme on l'observe chez le mouton domestique en raison du flux génétique avec le mouflon européen sauvage[54].
Le mélange soutenu entre différentes populations de chiens et de loups dans l'Ancien et le Nouveau Monde au cours des 10 000 dernières années au moins a brouillé les signatures génétiques et déjoué les efforts des chercheurs pour identifier les origines des chiens. Aucune des populations de loups modernes n'est liée aux loups du Pléistocène qui étaient les premiers a êtres domestiqués[55], et l'extinction des loups qui étaient les ancêtres directs des chiens a brouillé les efforts pour déterminer le moment et le lieu de la domestication des chiens[7].
Sélection positive
Charles Darwin a reconnu le petit nombre de traits qui différenciaient les espèces domestiques de leurs ancêtres sauvages. Il a également été le premier à reconnaître la différence entre l'élevage sélectif conscient dans lequel les humains sélectionnent directement les traits désirables, et la sélection inconsciente où les traits évoluent comme un sous-produit de la sélection naturelle ou de la sélection sur d'autres traits[2] - [3] - [4].
Par rapport à leurs homologues sauvages, les animaux domestiques présentent des variations dans la couleur du pelage et la morphologie craniofaciale, une taille réduite du cerveau, des oreilles tombantes et des changements dans le système endocrinien et leur cycle de reproduction. L'expérience du renard argenté domestiqué a démontré que la sélection pour la docilité en quelques générations peut entraîner une modification des traits comportementaux, morphologiques et physiologiques[22] - [28]. En plus de démontrer que les traits phénotypiques domestiques pouvaient survenir par la sélection d'un trait comportemental, et que les traits comportementaux domestiques pouvaient survenir par la sélection d'un trait phénotypique, ces expériences ont fourni un mécanisme pour expliquer comment le processus de domestication des animaux aurait pu commencer sans prévoyance humaine délibérée[28]. Dans les années 1980, un chercheur a utilisé un ensemble de marqueurs phénotypiques comportementaux, cognitifs et visibles, comme la couleur du pelage, pour produire des daims domestiques en quelques générations[28] - [56]. Des résultats similaires pour la docilité et la peur ont été trouvés pour le vison[57] et le quai japonais[58].
La différence génétique entre les populations domestiques et sauvages peut être encadrée par deux considérations. La première distingue les caractères de domestication présumés essentiels aux premiers stades de la domestication, et les caractères d'amélioration apparus depuis la scission entre populations sauvages et domestiques[5] - [6] - [7]. Les traits de domestication sont généralement fixés dans tous les domestiqués et ont été sélectionnés lors de l'épisode initial de domestication, alors que les traits d'amélioration ne sont présents que dans une proportion de domestiqués, bien qu'ils puissent être fixés dans des races individuelles ou des populations régionales[6] - [7] - [8]. Une deuxième question est de savoir si les traits associés au syndrome de domestication résultaient d'un relâchement de la sélection lorsque les animaux quittaient l'environnement sauvage ou d'une sélection positive résultant d'une préférence humaine intentionnelle et non intentionnelle. Certaines études génomiques récentes sur la base génétique des traits associés au syndrome de domestication ont mis en lumière ces deux problèmes[7].
Les généticiens ont identifié plus de 300 locus génétiques et 150 gènes associés à la variabilité de la couleur du pelage[28] - [59]. La connaissance des mutations associées aux différentes couleurs a permis une certaine corrélation entre le moment de l'apparition des couleurs de robe variables chez les chevaux et le moment de leur domestication[28] - [59]. D'autres études ont montré comment la sélection induite par l'homme est responsable de la variation allélique chez le porc[28] - [60]. Ensemble, ces informations suggèrent que, bien que la sélection naturelle ait maintenu la variation au minimum avant la domestication, les humains ont activement sélectionné de nouvelles couleurs de pelage dès leur apparition dans les populations gérées[28] - [35].
En 2015, une étude a examiné plus de 100 séquences du génome porcin pour déterminer leur processus de domestication. On a supposé que le processus de domestication avait été initié par l'homme, impliquait peu d'individus et reposait sur l'isolement reproductif entre les formes sauvages et domestiques, mais l'étude a révélé que l'hypothèse d'un isolement reproductif avec des goulots d'étranglement de la population n'était pas étayée. L'étude a indiqué que les porcs étaient domestiqués séparément en Asie occidentale et en Chine, les porcs d'Asie occidentale étant introduits en Europe où ils se sont croisés avec des sangliers. Un modèle qui correspondait aux données comprenait un mélange avec une population fantôme maintenant éteinte de cochons sauvages pendant le Pléistocène. L'étude a également révélé que malgré le rétrocroisement avec des porcs sauvages, les génomes des porcs domestiques présentent de fortes signatures de sélection au niveau des locus génétiques qui affectent le comportement et la morphologie. L'étude a conclu que la sélection humaine pour les traits domestiques a probablement contrecarré l'effet d'homogénéisation du flux génétique des sangliers et créé des îlots de domestication dans le génome. Le même processus peut également s'appliquer à d'autres animaux domestiques[19] - [61].
Contrairement à d'autres espèces domestiques qui ont été principalement sélectionnées pour des traits liés à la production, les chiens ont d'abord été sélectionnés pour leurs comportements[18] - [62]. En 2016, une étude a révélé qu'il n'y avait que 11 gènes fixes qui présentaient une variation entre les loups et les chiens. Il est peu probable que ces variations génétiques aient été le résultat d'une évolution naturelle et indiquent une sélection à la fois sur la morphologie et le comportement lors de la domestication du chien. Il a été démontré que ces gènes affectent la voie de synthèse des catécholamines, la majorité des gènes affectant la réponse de combat ou de fuite (c'est-à-dire la sélection pour la docilité) et le traitement émotionnel[18]. Les chiens montrent généralement moins de peur et d'agressivité que les loups[18] - [63]. Certains de ces gènes ont été associés à l'agressivité chez certaines races de chiens, indiquant leur importance à la fois dans la domestication initiale et plus tard dans la formation de la race[18].
Notes et références
- (en) Melinda A. Zeder, « Core questions in domestication research », Proceedings of the National Academy of Sciences, vol. 112, no 11, , p. 3191–3198 (ISSN 0027-8424 et 1091-6490, PMID 25713127, PMCID PMC4371924, DOI 10.1073/pnas.1501711112, lire en ligne, consulté le )
- (en) Charles Darwin, The Variation of Plants and Animals under Domestication, Londres, John Murray (OCLC 156100686)
- (en) Jared Diamond, Guns, Germs and Steel, Londres, Chatto and Windus, (ISBN 978-0-09-930278-0)
- (en) Greger Larson, Dolores R. Piperno, Robin G. Allaby et Michael D. Purugganan, « Current perspectives and the future of domestication studies », Proceedings of the National Academy of Sciences, vol. 111, no 17, , p. 6139–6146 (ISSN 0027-8424 et 1091-6490, PMID 24757054, PMCID PMC4035915, DOI 10.1073/pnas.1323964111, lire en ligne, consulté le )
- (en) Kenneth M. Olsen et Jonathan F. Wendel, « A Bountiful Harvest: Genomic Insights into Crop Domestication Phenotypes », Annual Review of Plant Biology, vol. 64, no 1, , p. 47–70 (ISSN 1543-5008 et 1545-2123, DOI 10.1146/annurev-arplant-050312-120048, lire en ligne, consulté le )
- (en) Andrew N. Doust, Lewis Lukens, Kenneth M. Olsen et Margarita Mauro-Herrera, « Beyond the single gene: How epistasis and gene-by-environment effects influence crop domestication », Proceedings of the National Academy of Sciences, vol. 111, no 17, , p. 6178–6183 (ISSN 0027-8424 et 1091-6490, PMID 24753598, PMCID PMC4035984, DOI 10.1073/pnas.1308940110, lire en ligne, consulté le )
- (en) Greger Larson et Dorian Q. Fuller, « The Evolution of Animal Domestication », Annual Review of Ecology, Evolution, and Systematics, vol. 45, no 1, , p. 115–136 (ISSN 1543-592X et 1545-2069, DOI 10.1146/annurev-ecolsys-110512-135813, lire en ligne, consulté le )
- (en) Rachel S. Meyer et Michael D. Purugganan, « Evolution of crop species: genetics of domestication and diversification », Nature Reviews Genetics, vol. 14, no 12, , p. 840–852 (ISSN 1471-0056 et 1471-0064, DOI 10.1038/nrg3605, lire en ligne, consulté le )
- (en) Edward Price, Principles and Applications of Domestic Animal Behaviour: An Introductory Text, Cambridge University Press, (ISBN 9781780640556)
- (en) Carlos A. Driscoll, David W. Macdonald et Stephen J. O'Brien, « From wild animals to domestic pets, an evolutionary view of domestication », Proceedings of the National Academy of Sciences, vol. 106, no supplement_1, , p. 9971–9978 (ISSN 0027-8424 et 1091-6490, PMID 19528637, PMCID PMC2702791, DOI 10.1073/pnas.0901586106, lire en ligne, consulté le )
- (en) Melinda A. Zeder, « The Domestication of Animals », Journal of Anthropological Research, vol. 68, no 2, , p. 161–190 (ISSN 0091-7710 et 2153-3806, DOI 10.3998/jar.0521004.0068.201, lire en ligne, consulté le )
- (en) E.B. Hale, "Domestication and the evolution of behavior". In Hafez, E. S. E. (ed.). The Behavior of Domestic Animals (2nd ed.), London, Bailliere, Tindall, and Cassell., , p. 22–42
- (en) Laurent Frantz, Erik Meijaard, Jaime Gongora et James Haile, « The Evolution of Suidae », Annual Review of Animal Biosciences, vol. 4, no 1, , p. 61–85 (ISSN 2165-8102 et 2165-8110, DOI 10.1146/annurev-animal-021815-111155, lire en ligne, consulté le )
- (en) Fiona B. Marshall, Keith Dobney, Tim Denham et José M. Capriles, « Evaluating the roles of directed breeding and gene flow in animal domestication », Proceedings of the National Academy of Sciences, vol. 111, no 17, , p. 6153–6158 (ISSN 0027-8424 et 1091-6490, PMID 24753599, PMCID PMC4035985, DOI 10.1073/pnas.1312984110, lire en ligne, consulté le )
- (en) Richard Blaustein, « Unraveling the Mysteries of Animal Domestication », BioScience, vol. 65, no 1, , p. 7–13 (ISSN 1525-3244 et 0006-3568, DOI 10.1093/biosci/biu201, lire en ligne, consulté le )
- (en) Greger Larson, Elinor K. Karlsson, Angela Perri et Matthew T. Webster, « Rethinking dog domestication by integrating genetics, archeology, and biogeography », Proceedings of the National Academy of Sciences, vol. 109, no 23, , p. 8878–8883 (ISSN 0027-8424 et 1091-6490, PMID 22615366, PMCID PMC3384140, DOI 10.1073/pnas.1203005109, lire en ligne, consulté le )
- (en) Angela Perri, « A wolf in dog's clothing: Initial dog domestication and Pleistocene wolf variation », Journal of Archaeological Science, vol. 68, , p. 1–4 (ISSN 0305-4403, DOI 10.1016/j.jas.2016.02.003, lire en ligne, consulté le )
- (en) Alex Cagan et Torsten Blass, « Identification of genomic variants putatively targeted by selection during dog domestication », BMC Evolutionary Biology, vol. 16, no 1, (ISSN 1471-2148, PMID 26754411, PMCID PMC4710014, DOI 10.1186/s12862-015-0579-7, lire en ligne, consulté le )
- (en) Laurent A F Frantz, Joshua G Schraiber, Ole Madsen et Hendrik-Jan Megens, « Evidence of long-term gene flow and selection during domestication from analyses of Eurasian wild and domestic pig genomes », Nature Genetics, vol. 47, no 10, , p. 1141–1148 (ISSN 1061-4036 et 1546-1718, DOI 10.1038/ng.3394, lire en ligne, consulté le )
- Elizabeth Pennisi, « The taming of the pig took some wild turns », Science, (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.aad1692, lire en ligne, consulté le )
- (en) Gillian P. McHugo, Michael J. Dover et David E. MacHugh, « Unlocking the origins and biology of domestic animals using ancient DNA and paleogenomics », BMC Biology, vol. 17, no 1, (ISSN 1741-7007, PMID 31791340, PMCID PMC6889691, DOI 10.1186/s12915-019-0724-7, lire en ligne, consulté le )
- (en) David E. MacHugh, Greger Larson et Ludovic Orlando, « Taming the Past: Ancient DNA and the Study of Animal Domestication », Annual Review of Animal Biosciences, vol. 5, no 1, , p. 329–351 (ISSN 2165-8102 et 2165-8110, DOI 10.1146/annurev-animal-022516-022747, lire en ligne, consulté le )
- Barker, G. (2006). The Agricultural Revolution in Prehistory: Why Did Foragers Become Farmers?. Oxford University Press.
- (en) David E. MacHugh, Greger Larson et Ludovic Orlando, « Taming the Past: Ancient DNA and the Study of Animal Domestication », Annual Review of Animal Biosciences, vol. 5, no 1, , p. 329–351 (ISSN 2165-8102 et 2165-8110, DOI 10.1146/annurev-animal-022516-022747, lire en ligne, consulté le )
- Fuller DQ, Willcox G, Allaby RG. 2011. Cultivation and domestication had multiple origins: arguments against the core area hypothesis for the origins of agriculture in the Near East. World Archaeol. 43:628–52
- Melinda A. Zeder 2006. Archaeological approaches to documenting animal domestication. In Documenting Domestication: New Genetic and Archaeological Paradigms, ed. M.A. Zeder, D.G Bradley, E Emshwiller, B.D Smith, pp. 209–27. Berkeley: Univ. Calif. Press
- Valclav Smil, 2011, Harvesting the Biosphere:The Human Impact, Population and Development Review 37(4): 613–636, Table 2.
- (en) Greger Larson et Joachim Burger, « A population genetics view of animal domestication », Trends in Genetics, vol. 29, no 4, , p. 197–205 (DOI 10.1016/j.tig.2013.01.003, lire en ligne, consulté le )
- Melinda A. Zeder. 2006. Archaeological approaches to documenting animal domestication. In Documenting Domestication: New Genetic and Archaeological Paradigms, ed. M.A. Zeder, D.G Bradley, E Emshwiller, B.D Smith, pp. 209–27. Berkeley: University of California Press.
- Kruska, D. 1988. "Mammalian domestication and its effect on brain structure and behavior," in Intelligence and evolutionary biology. Edited by H. J. Jerison and I. Jerison, pp. 211–50. New York: Springer-Verlag
- (en) Lyudmila Trut, « Early Canid Domestication: The Farm-Fox Experiment », American Scientist, vol. 87, no 2, , p. 160 (ISSN 0003-0996 et 1545-2786, DOI 10.1511/1999.2.160, lire en ligne, consulté le )
- (en) Lyudmila Trut, Irina Oskina et Anastasiya Kharlamova, « Animal evolution during domestication: the domesticated fox as a model », BioEssays, vol. 31, no 3, , p. 349–360 (PMID 19260016, PMCID PMC2763232, DOI 10.1002/bies.200800070, lire en ligne, consulté le )
- (en) Adam S Wilkins, Richard W Wrangham et W Tecumseh Fitch, « The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and Genetics », Genetics, vol. 197, no 3, , p. 795–808 (ISSN 1943-2631, PMID 25024034, PMCID PMC4096361, DOI 10.1534/genetics.114.165423, lire en ligne, consulté le )
- (en) Dominic Wright, « Article Commentary: The Genetic Architecture of Domestication in Animals », Bioinformatics and Biology Insights, vol. 9S4, , BBI.S28902 (ISSN 1177-9322 et 1177-9322, PMID 26512200, PMCID PMC4603525, DOI 10.4137/BBI.S28902, lire en ligne, consulté le )
- Hemmer, H. (1990). Domestication: The Decline of Environmental Appreciation. Cambridge University Press.
- (en) J.D.S. Birks et A.C. Kitchener, 1999. The distribution and status of the polecat Mustela putorius in Britain in the 1990s. Vincent Wildlife Trust, Londres.
- (de) W. Schultz, 1969, « Zur kenntnis des hallstromhundes (Canis hallstromi, Troughton 1957) », Zoologischer Anzeiger, vol. 183, p. 42–72.
- (en) L. Boitani et P. Ciucci, « Comparative social ecology of feral dogs and wolves », Ethology Ecology & Evolution, vol. 7, no 1, , p. 49–72 (ISSN 0394-9370 et 1828-7131, DOI 10.1080/08927014.1995.9522969, lire en ligne, consulté le )
- (en) Greger Larson et Joachim Burger, « A population genetics view of animal domestication », Trends in Genetics, vol. 29, no 4, , p. 197–205 (DOI 10.1016/j.tig.2013.01.003, lire en ligne, consulté le )
- (en) Jean-Denis Vigne, « The origins of animal domestication and husbandry: A major change in the history of humanity and the biosphere », Comptes Rendus Biologies, vol. 334, no 3, , p. 171–181 (DOI 10.1016/j.crvi.2010.12.009, lire en ligne, consulté le )
- Crockford, S. J. (2000). "A commentary on dog evolution: Regional variation, breed development and hybridization with wolves". In Crockford, S. (ed.). Dogs through Time: An Archaeological Perspective. BAR International Series 889. Oxford: Archaeopress. pp. 11–20. ISBN 978-1841710891.
- Coppinger, Raymond; Coppinger, Laura (2001). Dogs: A Startling New Understanding of Canine Origin, Behavior & Evolution. Scribner. ISBN 978-0684855301.
- Russell, N. (2012). Social Zooarchaeology: Humans and Animals in Prehistory. Cambridge University Press. ISBN 978-0-521-14311-0.
- Evan K. Irving-Pease, Hannah Ryan, Alexandra Jamieson et Evangelos A. Dimopoulos, « Paleogenomics of Animal Domestication », dans Paleogenomics, Springer International Publishing, (ISBN 978-3-030-04752-8, DOI 10.1007/13836_2018_55, lire en ligne), p. 225–272
- (en) Ann Gibbons, « How an ancient pope helped make chickens fat », Science, (DOI 10.1126/science.aah7308, lire en ligne, consulté le )
- (en) Jared Diamond, « Evolution, consequences and future of plant and animal domestication », Nature, vol. 418, no 6898, , p. 700–707 (ISSN 1476-4687, DOI 10.1038/nature01019, lire en ligne, consulté le )
- (en) Birgitta Kimura, Fiona Marshall, Albano Beja-Pereira et Connie Mulligan, « Donkey Domestication », African Archaeological Review, vol. 30, no 1, , p. 83–95 (ISSN 0263-0338 et 1572-9842, DOI 10.1007/s10437-012-9126-8, lire en ligne, consulté le )
- (en) Mathias Currat, Manuel Ruedi, Rmy J. Petit et Laurent Excoffier, « THE HIDDEN SIDE OF INVASIONS: MASSIVE INTROGRESSION BY LOCAL GENES », Evolution, , ???–??? (DOI 10.1111/j.1558-5646.2008.00413.x, lire en ligne, consulté le )
- (en) Jonas Eriksson, Greger Larson, Ulrika Gunnarsson et Bertrand Bed'hom, « Identification of the Yellow Skin Gene Reveals a Hybrid Origin of the Domestic Chicken », PLoS Genetics, vol. 4, no 2, , e1000010 (ISSN 1553-7404, PMID 18454198, PMCID PMC2265484, DOI 10.1371/journal.pgen.1000010, lire en ligne, consulté le )
- (en) Olivier Hanotte, Daniel G. Bradley, Joel W. Ochieng et Yasmin Verjee, « African Pastoralism: Genetic Imprints of Origins and Migrations », Science, vol. 296, no 5566, , p. 336–339 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.1069878, lire en ligne, consulté le )
- (en) Edward L. C. Verkaar, Isaäc J. Nijman, Maurice Beeke et Eline Hanekamp, « Maternal and Paternal Lineages in Cross-Breeding Bovine Species. Has Wisent a Hybrid Origin? », Molecular Biology and Evolution, vol. 21, no 7, , p. 1165–1170 (ISSN 1537-1719 et 0737-4038, DOI 10.1093/molbev/msh064, lire en ligne, consulté le )
- (en) J. Jordana, P.M. Pares et A. Sanchez, « Analysis of genetic relationships in horse breeds », Journal of Equine Veterinary Science, vol. 15, no 7, , p. 320–328 (DOI 10.1016/S0737-0806(06)81738-7, lire en ligne, consulté le )
- (en) Brock A. Harpur, Shermineh Minaei, Clement F. Kent et Amro Zayed, « Management increases genetic diversity of honey bees via admixture: GENETIC DIVERSITY IN THE HONEY BEE », Molecular Ecology, vol. 21, no 18, , p. 4414–4421 (DOI 10.1111/j.1365-294X.2012.05614.x, lire en ligne, consulté le )
- (en) Mario Barbato, Frank Hailer, Pablo Orozco-terWengel et James Kijas, « Genomic signatures of adaptive introgression from European mouflon into domestic sheep », Scientific Reports, vol. 7, no 1, , p. 7623 (ISSN 2045-2322, PMID 28790322, PMCID PMC5548776, DOI 10.1038/s41598-017-07382-7, lire en ligne, consulté le )
- (en) Adam H. Freedman, Ilan Gronau, Rena M. Schweizer et Diego Ortega-Del Vecchyo, « Genome Sequencing Highlights the Dynamic Early History of Dogs », PLoS Genetics, vol. 10, no 1, , e1004016 (ISSN 1553-7404, PMID 24453982, PMCID PMC3894170, DOI 10.1371/journal.pgen.1004016, lire en ligne, consulté le )
- Hemmer, H. (2005). "Neumuhle-Riswicker Hirsche: Erste planma¨ßige Zucht einer neuen Nutztierform". Naturwissenschaftliche Rundschau. 58: 255–261.
- (en) Jens Malmkvist et Steffen W. Hansen, « Generalization of fear in farm mink, Mustela vison, genetically selected for behaviour towards humans », Animal Behaviour, vol. 64, no 3, , p. 487–501 (DOI 10.1006/anbe.2002.3058, lire en ligne, consulté le )
- (en) R.Bryan Jones, Daniel G. Satterlee et Henry L. Marks, « Fear-related behaviour in Japanese quail divergently selected for body weight », Applied Animal Behaviour Science, vol. 52, nos 1-2, , p. 87–98 (DOI 10.1016/S0168-1591(96)01146-X, lire en ligne, consulté le )
- (en) Arne Ludwig, Melanie Pruvost, Monika Reissmann et Norbert Benecke, « Coat Color Variation at the Beginning of Horse Domestication », Science, vol. 324, no 5926, , p. 485–485 (ISSN 0036-8075 et 1095-9203, PMID 19390039, PMCID PMC5102060, DOI 10.1126/science.1172750, lire en ligne, consulté le )
- (en) Meiying Fang, Greger Larson, Helena Soares Ribeiro et Ning Li, « Contrasting Mode of Evolution at a Coat Color Locus in Wild and Domestic Pigs », PLoS Genetics, vol. 5, no 1, , e1000341 (ISSN 1553-7404, PMID 19148282, PMCID PMC2613536, DOI 10.1371/journal.pgen.1000341, lire en ligne, consulté le )
- Elizabeth Pennisi, « The taming of the pig took some wild turns », Science, (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.aad1692, lire en ligne, consulté le )
- Serpell, J.; Duffy, D. (2014). "Dog Breeds and Their Behavior". Domestic Dog Cognition and Behavior. Berlin & Heidelberg: Springer.
- Coppinger, R.; Schneider, R. (1995). "Evolution of working dogs". The Domestic Dog: Its Evolution, Behaviour and Interactions with People. Cambridge University Press. ISBN 9780521425377.