Codage neuronal
Le codage neuronal désigne, en neurosciences, la relation hypothétique entre le stimulus et les réponses neuronales individuelles ou globales[1]. C'est une théorie sur l'activité électrique du système nerveux[2], selon laquelle les informations, par exemple sensorielles, numériques ou analogiques, sont représentées dans le cerveau par des réseaux de neurones[3]. Le codage neuronal est lié aux concepts du souvenir, de l'association et de la mémoire sensorielle.
Vue d'ensemble
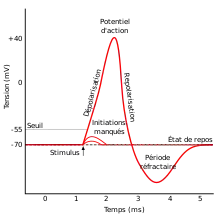
Les neurones propagent des signaux à grande vitesse à travers le corps, sous la forme d'impulsions électriques appelés potentiels d'action : l'évènement court de l'augmentation suivi de la chute du potentiel électrique de la neurone génère des pointes de tension (ou trains de pics) pouvant se déplacer le long des axones. En réaction à des stimulus sensoriels externes (comme la lumière, le son, le goût, l'odorat, le toucher), les neurones modifient leur activité et déclenchent différentes séquences de potentiels d'action. Ces informations sur le stimulus sont alors codées en un schéma de potentiels d’action, transmis au cerveau.
Les séquences de potentiels d'action ne sont pas des évènements identiques : leur durée, leur amplitude et leur forme peuvent varier. L'objet des études sur le codage neuronal est la représentation des attributs de stimulus (par exemple, l'intensité lumineuse ou sonore et les actions motrices par les pics de neurones. Si chaque potentiel d'action dure environ une milliseconde, une séquence de potentiels d'action (ou train de pics) peut être considérée comme série d'événements ponctuels séparés par des intervalles de longueur variable[4] - [5]. Pour la description et l'analyse du déclenchement neuronal, des méthodes statistiques et des méthodes de la théorie des probabilités et des processus ponctuels stochastiques sont appliquées.
Avec le développement de techniques d'enregistrement et de décodage en neurosciences, les chercheurs arrivent à déchiffrer le code neuronal. Un premier aperçu de ce codage en temps réel révèle que l'hippocampe joue un rôle central dans la mémoire[6] - [7] - [8]. En 2020, plusieurs projets de décodage cérébral sont en développement[9] - [10].
Encodage et décodage
Deux points de vue opposés dirigent l'étude du lien entre stimulus et réponse neuronale : encodage et décodage.
- L'encodage neuronal se réfère au chemin du stimulus vers la réponse. L'objectif principal est de comprendre comment les neurones répondent aux stimulus et de construire des modèles visant la prédiction de réponses à d'autres types de stimulus.
- Le décodage neuronal se réfère au chemin de la réponse vers le stimulus. Le défi consiste alors à reconstruire un stimulus, ou certains de ses aspects, à partir des séquences de pics qui le représentent.
Hypothèses sur les schémas de codage
Une séquence ou un train de pics peut coder des informations basées selon différents types de schémas. Cette variation et le manque de savoir sur le sujet provoque de nombreux débats au sein de la communauté neuroscientifique, d'où plusieurs théories hypothétiques.
On avance que le "taux de déclenchement neuronal" (le nombre moyen de pics par unité de temps) détermine la force motrice, par exemple, l'énergie avec laquelle un muscle innervé est contracté. À l’inverse, on propose que le "code temporel complexe" est basé sur le minutage précis de pics uniques, qui peuvent être verrouillés par un stimulus externe (comme dans le système visuel et auditif) ou être générés intrinsèquement par les circuits neuronaux[11] - [12].
La théorie de la neuroélectrodynamique considère tous les schémas de codage comme des épiphénomènes liés aux changements moléculaires, où la distribution spatiale des champs électriques dans les neurones résulte du large spectre électromagnétique des potentiels d'action[13] - [14] - [15] - [16] - [17].
Taux de déclenchement
Le modèle de codage de la fréquence suppose que l'information sur un signal est contenue dans la fréquence de tir des pointes. Il est parfois appelé codage de fréquence, bien qu'à proprement parler, la fréquence des événements discrets ne soit pas une fréquence mais un tempo. Ainsi, appeler ce modèle un code tempo serait physiquement correct.
Il est apparu après des expériences menées par ED Adrian et Y Zotterman en 1926[18]. Dans cette expérience simple, différents poids étaient suspendus à un muscle. Lorsque le poids du stimulus augmentait, le nombre de pics enregistrés à partir des nerfs sensitifs innervant le muscle augmentait également. Les auteurs en ont conclu que les potentiels d'action étaient des événements discrets et que leur rythme, plutôt que des paramètres individuels, constituait la base de la communication neuronale. Au cours des décennies suivantes, la mesure des taux de tir est devenue un outil standard pour décrire les propriétés de tous les types de neurones, en partie en raison de la facilité relative de mesurer les taux de manière expérimentale. Cependant, cette approche néglige toutes les informations éventuellement contenues dans le timing exact des pics et des intervalles inter-pics ainsi que les paramètres internes de chaque potentiel d'action. Ces dernières années, de plus en plus de preuves expérimentales ont suggéré qu'un concept de taux d'allumage simple basé sur la moyenne temporelle pourrait être trop simpliste pour décrire l'activité cérébrale[4]. Même au niveau périphérique (capteurs et effecteurs), le taux d'allumage augmente de façon non linéaire avec l'intensité du stimulus[19]. Il n'y a pas de lien direct entre la fréquence des pics et le signal. En outre, la séquence des potentiels d'action générés par un stimulus donné varie d'un essai à l'autre, de sorte que les réponses neuronales sont généralement traitées de manière statistique ou probabiliste. Même le terme "taux de tir" a plusieurs définitions, qui font référence à différentes procédures de calcul de la moyenne, comme une moyenne dans le temps ou une moyenne sur plusieurs répétitions d'une expérience.
Codage temporel
Les modèles de code temporel supposent que la synchronisation précise des pointes et des intervalles entre pointes est porteuse d'information[20]. De plus en plus de preuves confirment cette hypothèse[21] - [22] - [23] - [24] - [25] - [26]
Les modèles de codage du taux suggèrent que les irrégularités du tir neuronal sont du bruit et en font la moyenne. Le codage temporel fournit une autre explication du "bruit", suggérant qu'il code en fait des informations et affecte le traitement neuronal[27]. Pour modéliser cette idée, symboles binaires peut être utilisé pour marquer les pics : 1 pour un pic, 0 pour l'absence de pic. Le codage temporel permet à la séquence 000111000111 de signifier quelque chose de différent de 001100110011, même si le taux de tir moyen est le même pour les deux séquences[28]. Ainsi, le modèle peut être appelé le code numérique.
Les fonctions du cerveau sont plus précises dans le temps que ne le permet le codage du taux. En outre, les réponses à des stimuli similaires sont suffisamment différentes pour suggérer que les schémas distincts de pics contiennent un volume d'informations plus important que celui qu'il est possible d'inclure dans un code de taux[29].
La structure temporelle d'un train de pointes évoqué par un stimulus est déterminée à la fois par la dynamique du stimulus et par la nature du processus d'encodage neuronal. Les stimuli qui changent rapidement ont tendance à générer des pointes précisément synchronisées[30]. Le codage temporel utilise des caractéristiques de l'activité de stimulation qui ne peuvent pas être décrites par le taux de tir. Par exemple, le temps jusqu'au premier pic après l'apparition du stimulus, la phase du tir par rapport aux oscillations de fond, les caractéristiques basées sur le second et les plus hauts moments statistiques de la distribution de probabilité d'intervalle, le caractère aléatoire des pics, ou les groupes de pics précisément chronométrés (modèles temporels) sont des candidats pour les codes temporels[31]. Comme il n'y a pas de référence temporelle absolue dans le système nerveux, l'information est transportée soit en termes de synchronisation relative des pics dans une population de neurones (modèles temporels), soit par rapport à une oscillation cérébrale continue. (phase de tir)[25] - [4]. L'un des mécanismes possibles du code temporel est que les pics survenant à des phases spécifiques d'un cycle oscillatoire sont plus efficaces pour dépolariser la neurone post-synaptique[32]. Dans le codage temporel, l'apprentissage peut être expliqué par des modifications du délai synaptique dépendant de l'activité[33]. Les modifications peuvent elles-mêmes dépendre des modèles de synchronisation des spikes (codage temporel), c'est-à-dire peut-être un cas particulier de plasticité dépendante du rythme des pointes[34].
Pour des stimuli très brefs, le taux d'excitation maximal d'un neurone peut ne pas être assez rapide pour produire plus d'un seul pic. En raison de la densité de l'information contenue dans ce pic unique, il semblerait que le moment du pic lui-même doive transmettre plus d'informations que le simple taux moyen de potentiels d'action sur une période donnée. Ce modèle est particulièrement important pour la localisation du son, qui se produit dans le cerveau à l'échelle de quelques millisecondes. Le cerveau doit obtenir une grande quantité d'informations sur la base d'une réponse neuronale relativement courte. De plus, si des taux de tir faibles, de l'ordre de dix pics par seconde, doivent être distingués d'un codage de taux arbitrairement proche pour des stimuli différents, alors un neurone essayant de discriminer ces deux stimuli peut avoir besoin d'attendre une seconde ou plus pour accumuler suffisamment d'informations. Ceci n'est pas cohérent avec les nombreux organismes qui sont capables de distinguer des stimuli dans un laps de temps de quelques millisecondes ou moins[28].
Pour rendre compte de l'encodage rapide des stimuli visuels, il a été suggéré que les neurones de la rétine encodent l'information visuelle dans le temps de latence entre le début du stimulus et le premier potentiel d'action, également appelé latence au premier pic ou temps au premier pic[35]. Ce type de codage temporel a également été démontré dans le système auditif et somato-sensoriel. Le principal inconvénient d'un tel schéma de codage est sa sensibilité aux fluctuations neuronales intrinsèques[36]. Dans le cortex visuel primaire de macaques, le moment du premier pic par rapport au début du stimulus s'est avéré fournir plus d'informations que l'intervalle entre les pics. Cependant, l'intervalle entre les pointes pourrait être utilisé pour coder des informations supplémentaires, ce qui est particulièrement important lorsque le taux de pointes atteint sa limite, comme dans les situations à fort contraste. Pour cette raison, le codage temporel pourrait jouer un rôle dans le codage de bords définis plutôt que de transitions graduelles[37].
Comme pour le système visuel, dans les cellules mitrales/tuftées du bulbe olfactif des souris, la latence du premier pic par rapport au début d'une action de reniflement semble encoder une grande partie de l'information sur une odeur. Cette stratégie d'utilisation de la latence des pics permet une identification et une réaction rapides à une substance odorante. En outre, certaines cellules mitrales/tuftées présentent des schémas de tir spécifiques pour des odorants donnés. Dans le même ordre d'idées, des expériences réalisées avec le système olfactif de lapins ont montré des schémas distincts qui étaient corrélés à différents sous-ensembles d'odorants, et un résultat similaire a été obtenu lors d'expériences avec le système olfactif de criquets[28].
Le système gustatif des mammifères est utile pour l'étude du codage temporel en raison de ses stimuli assez distincts et des réponses facilement discernables de l'organisme[38]. L'information codée temporellement peut aider un organisme à discriminer entre différents agents gustatifs de la même catégorie (sucré, amer, acide, salé, umami) qui suscitent des réponses très similaires en termes de nombre de pics. La composante temporelle du schéma déclenché par chaque agent gustatif peut être utilisée pour déterminer son identité (par exemple, la différence entre deux agents gustatifs amers, tels que la quinine et le denatonium). De cette façon, le système gustatif peut utiliser à la fois le codage de la vitesse et le codage temporel - la vitesse pour le type de tastant de base, le temporel pour une différenciation plus spécifique[39]. La recherche sur le système gustatif des mammifères a montré qu'il y a une abondance d'informations présentes dans les modèles temporels à travers les populations de neurones, et ces informations sont différentes de celles qui sont déterminées par les schémas de codage du taux. Dans des études portant sur la partie corticale antérieure du cerveau chez les primates, des modèles précis avec des échelles de temps courtes de quelques millisecondes seulement ont été trouvés dans de petites populations de neurones en corrélation avec certains comportements. Cependant, peu d'informations ont pu être déterminées à partir de ces schémas. L'une des explications est que l'activité des cortical neurones ne correspond pas linéairement à la dynamique des paramètres du signal entrant car la chaîne technologique est constituée de convertisseurs de signaux primaires (capteurs), de modulateurs (structures sous-corticales) et d'intégrateurs (populations corticales) qui ne reflètent pas le signal mais le transduisent et créent des représentations[40].
L'hypothèse selon laquelle le code neuronal est binaire (les pointes et les intervalles inter-pointes étant des 1 et des 0) augmente considérablement la capacité du code et rend le modèle plus plausible. Mais la même question se pose de corréler la capacité d'information du code et la vitesse réelle du cerveau, qui parvient à coder un signal multiparamétrique complexe en un ou deux spikes. Le cerveau n'a pas le temps de construire une longue chaîne binaire qui pourrait contenir toutes les informations. En cela, il est fondamentalement différent des systèmes numériques artificiels. Malgré l'énorme vitesse de leurs processeurs, qui sont supérieurs de plusieurs ordres de grandeur aux fréquences du cerveau, ils ne peuvent l'égaler en termes de performances, de rapidité et d'efficacité énergétique. Le problème est qu'ils doivent traiter de longues chaînes de codes binaires. Le cerveau doit utiliser une capacité supplémentaire dans son code.
En outre, la question de l'horloge du système se pose à nouveau. Deux zéros du code représentent une pause deux fois plus longue qu'un zéro. Mais comment déterminer que la pause inter-pic signifie deux zéros ou un seul si l'on ne connaît pas l'échelle de temps du système étudié ? La mesure de la pause par une horloge externe donne beaucoup de données, mais ne dit rien sur le nombre de zéros dans cette pause particulière et sur leur relation avec les unités de pointes. En d'autres termes, nous ne pouvons pas déterminer si l'activité des neurones signifie 0001 ou 001. Pour une véritable analyse qualitative, il est nécessaire de normaliser les données du système par son propre temps. Nous pouvons alors exprimer notre analyse dans n'importe quelle unité de mesure. Trouver cette fréquence fondamentale comme base de normalisation est probablement d'une importance capitale lorsqu'on essaie de déchiffrer le code du cerveau, quel que soit le modèle de code que l'on teste, puisque le paramètre temps demeure de toute façon.
Ultramicro-codage sans pointe (pour une intelligence avancée)
Quels que soient les mérites et l'omniprésence de la signalisation « Potentiel d’action / Synaptique » (« spike » ou « pointe ») et de son codage, elle semble incapable d'offrir une explication plausible de « l'intelligence supérieure » telle que la pensée abstraite humaine ; par exemple. voir[41] - [42] D'où une recherche d'une alternative capable de numérique fiable performance, mais le seul candidat plausible semblait être l'utilisation d'ARN 'de réserve' (ne pas impliqué dans la codage des protéines d'où "ARNnc"). Ce ARNnc offrirait le codage statique "écrit". De tels sites ultramicro ne pourraient pas systématiquement communiquer entre eux en utilisant des potentiels d'action, mais ils devraient presque certainement utiliser infra-rouge ou des longueurs d'onde optiques proches. De telles longueurs d'onde s'adapteraient commodément aux diamètres des fibres nerveuses myélinisées — ici considérées comme des câbles coaxiaux — offrant ainsi un deuxième système de signalisation rapide (avec des propriétés significativement différentes) fonctionnant simultanément avec le système traditionnel, sur les mêmes axones chaque fois que nécessaire[43] - [44] (traduit par[45]), et[46].
Même si nous l'acceptons comme vrai, une telle activité est pour la plupart inobservable — pas directement observable pour des raisons pratiques — donc la mesure dans laquelle on devrait accepter ce modèle dépend de sa philosophie des sciences. Le modèle est basé sur une quantité considérable de preuves interdisciplinaires qui se soutiennent mutuellement, donc le réalisme scientifique devrait probablement l'accepter (tout comme il le fait pour les trous noirs ou neutrinos invisibles), au moins jusqu'à ce qu'une certaine réfutation "observée" surgit — tandis que l'on pourrait s'attendre à ce que l'instrumentalisme mélange l'incrédulité avec une volonté d'utiliser simplement le modèle comme utile dans la pratique, étant donné qu'il répond à plusieurs mystères.
Mais en plus, il y a deux éléments mineurs de preuve "directe" sous la forme de prédictions réalisées : (i) (plus un espoir qu'une prédiction) qu'il y aurait suffisamment d'ARN disponible — un doute qui a été dissipé lorsque Mattick[47] ont révélé que (chez l'homme) seulement environ 3% de l'ARN était utilisé pour la fabrication de protéines, "donc 97% étaient disponibles pour d'autres tâches". (ii) La faisabilité de la sous-hypothèse du câble coaxial a été justifiée par des expériences montrant que l'infrarouge et d'autres fréquences lumineuses peuvent être transmises via les axones[48] - [49]. Ce non-spike mode est envisagé comme fonctionnant "exclusivement dans le cerveau proprement dit" — en tant que mécanismes de pensée avancée (chez les vertébrés supérieurs) — laissant les signaux conventionnels de "pointe" faire toute l'intercommunication avec le monde extérieur, et faire d'autres tâches de routine, y compris Entretien hebbien.
Étonnamment cependant, il a été suggéré qu'un mode similaire aurait évolué indépendamment chez les insectes (expliquant ainsi leurs capacités de performance extraordinaires malgré leur petit cerveau). En effet, comme il existe un cas où les épines et les antennes des papillons de nuit, etc. peuvent recevoir des signaux infrarouges directement de l'environnement[50], (revue[51]), il existe donc une possibilité supplémentaire qu'il puisse parfois y avoir une alimentation dédiée de ces signaux directement dans le système nerveux de l'insecte (sans les mécanismes sensoriels de « pointe » habituellement attendus). Ce n'est qu'une conjecture à ce stade, mais cela pourrait offrir la possibilité d'une expérimentation facile et économique.
Encore un autre mode de signal sans pointe : Il existe également des preuves indirectes d'un troisième mode de signal pour l'axone ! Ce mode est beaucoup plus lent mais capable de transporter des documents beaucoup plus volumineux sous la forme de schémas d'ARNnc déjà formatés du codage statique mentionné ci-dessus — transportés en tant que transport axonal par kinésine dans l'axone[45] - [44] (tout comme le transport connu de ARNm, avec lequel il peut avoir été confondu dans des études en laboratoire).
Voir également
- Réseau de neurones artificiels
- Auto-encodeur
- Modèles du neurone biologique
- Binding problem
- Cognitive map
- Deep learning
- Feature integration theory
- Théorie du neurone grand-mère
- Models of neural computation
- Neural correlate
- Neural decoding
- Neural oscillation
- Sparse distributed memory
- Quantification vectorielle
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Neural coding » (voir la liste des auteurs).
- (zh-yue) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en cantonais intitulé « 神經編碼 » (voir la liste des auteurs).
- (en) « Multiple neural spike train data analysis: state-of-the-art and future challenges », Nat. Neurosci., vol. 7, no 5, , p. 456–61 (PMID 15114358, DOI 10.1038/nn1228).
- (en) Johnson, « Neural coding », Neuron, vol. 26, no 3, , p. 563–566 (ISSN 0896-6273, PMID 10896153, DOI 10.1016/S0896-6273(00)81193-9).
- (en) S.J. Thorpe, Parallel processing in neural systems and computers, North-Holland, , 91–94 p., PDF (ISBN 978-0-444-88390-2, lire en ligne), « Spike arrival times: A highly efficient coding scheme for neural networks ».
- (en) « Neuronal variability: noise or part of the signal? », Nat. Rev. Neurosci., vol. 6, no 5, , p. 389–97 (PMID 15861181, DOI 10.1038/nrn1668).
- (en) Wulfram Gerstner et Werner M. Kistler, Spiking Neuron Models : Single Neurons, Populations, Plasticity, Cambridge University Press, , 480 p. (ISBN 978-0-521-89079-3, lire en ligne).
- (en) The Memory Code. http://www.scientificamerican.com/article/the-memory-code/
- (en) Chen, Wang et Tsien, « Neural population-level memory traces in the mouse hippocampus », PLOS ONE, vol. 4, no 12, , e8256 (PMID 20016843, PMCID 2788416, DOI 10.1371/journal.pone.0008256, Bibcode 2009PLoSO...4.8256C).
- (en) Zhang, Chen, Kuang et Tsien, « Mapping and deciphering neural codes of NMDA receptor-dependent fear memory engrams in the hippocampus », PLOS ONE, vol. 8, no 11, , e79454 (PMID 24302990, PMCID 3841182, DOI 10.1371/journal.pone.0079454, Bibcode 2013PLoSO...879454Z).
- (en) Brain Decoding Project. http://braindecodingproject.org/
- (en) The Simons Collaboration on the Global Brain. https://www.simonsfoundation.org/life-sciences/simons-collaboration-global-brain/
- (en) Burcas G.T & Albright T.D. Gauging sensory representations in the brain. http://www.vcl.salk.edu/Publications/PDF/Buracas_Albright_1999_TINS.pdf
- (en) « Neural codes: firing rates and beyond », Proc. Natl. Acad. Sci. U.S.A., vol. 94, no 24, , p. 12740–1 (PMID 9398065, PMCID 34168, DOI 10.1073/pnas.94.24.12740, Bibcode 1997PNAS...9412740G).
- (en) Aur D., Jog, MS., 2010 Neuroelectrodynamics: Understanding the brain language, IOS Press, 2010, DOI 10.3233/978-1-60750-473-3-i.
- (en) Aur, Connolly et Jog, « Computing spike directivity with tetrodes », J. Neurosci., vol. 149, no 1, , p. 57–63 (PMID 15978667, DOI 10.1016/j.jneumeth.2005.05.006).
- (en) Aur et Jog, « Reading the Neural Code: What do Spikes Mean for Behavior? », Nature Precedings, (DOI 10.1038/npre.2007.61.1).
- (en) Fraser et Frey, « Electromagnetic emission at micron wavelengths from active nerves », Biophysical Journal, vol. 8, no 6, , p. 731–734 (PMID 5699805, PMCID 1367349, DOI 10.1016/s0006-3495(68)86517-8, Bibcode 1968BpJ.....8..731F).
- (en) Aur, « A comparative analysis of integrating visual information in local neuronal ensembles », Journal of Neuroscience Methods, vol. 207, no 1, , p. 23–30 (PMID 22480985, PMCID 3636996, DOI 10.1016/j.jneumeth.2012.03.008).
- Adrian ED, Zotterman Y, « Les impulsions produites par les terminaisons nerveuses sensorielles : Part II : The response of a single end organ. », J Physiol, vol. 61, no 2, , p. 151-171 (PMID 16993780, PMCID 1514782, DOI 10.1113/jphysiol.1926.sp002281)
- E. Kandel, J. Schwartz et T. M. Jessel, Principles of Neural Science, Elsevier, , 3rd éd. (ISBN 978-0444015624, lire en ligne)
- Peter Dayan et L. F. Abbott, Neurosciences théoriques : Computational and Mathematical Modeling of Neural Systems, Massachusetts Institute of Technology Press, (ISBN 978-0-262-04199-7, lire en ligne)
- (en) T. Gollisch et M. Meister, « Rapid Neural Coding in the Retina with Relative Spike Latencies », Science, vol. 319, no 5866, , p. 1108–1111 (ISSN 0036-8075, PMID 18292344, DOI 10.1126/science.1149639, Bibcode 2008Sci...319.1108G, S2CID 1032537, lire en ligne)
- Forrest MD, « La dynamique calcique intracellulaire permet à un modèle de neurone de Purkinje d'effectuer des calculs de bascule et de gain sur ses entrées. », Frontiers in Computational Neuroscience, vol. 8, , p. 86 (PMID 25191262, PMCID 4138505, DOI 10.3389/fncom.2014 .00086
 )
) - Forrest MD, « La pompe sodium-potassium est un élément de traitement de l'information dans le calcul cérébral », Frontiers in Physiology, vol. 5, no 472, , p. 472 (PMID 25566080, PMCID 4274886, DOI 10.3389/fphys.2014 .00472 )
- Singh & Levy, " A consensus layer V pyramidal neuron can sustain interpulse-interval coding ", PLoS ONE, 2017
- S.J. Thorpe, Parallel processing in neural systems and computers, North-Holland, , 91-94 p., PDF (ISBN 978-0-444-88390-2, lire en ligne), « Temps d'arrivée des pics : A highly efficient coding scheme for neural networks »
- Butts DA, Weng C, Jin J, etal, « Précision temporelle dans le code neural et les échelles de temps de la vision naturelle », Nature, vol. 449, no 7158, , p. 92-5 (PMID 17805296, DOI 10. 1038/nature06105, Bibcode 2007Natur.449 ...92B, S2CID 4402057)
- J. Leo van Hemmen, TJ Sejnowski. 23 Problèmes dans la neuroscience des systèmes. Oxford Univ. Press, 2006. p.143-158.
- F Theunissen et JP Miller, « Temporal Encoding in Nervous Systems : A Rigorous Definition », Journal of Computational Neuroscience, vol. 2, no 2, , p. 149-162 (PMID 8521284, DOI 10.1007/bf00961885, S2CID 206786736)
- Charles, Anthony Zador, Stevens, « L'énigme du cerveau », sur © Current Biology 1995, Vol 5 No 12 (consulté le )
- (en) Renaud Jolivet, Alexander Rauch, Hans-Rudolf Lüscher et Wulfram Gerstner, « Predicting spike timing of neocortical pyramidal neurons by simple threshold models », Journal of Computational Neuroscience, vol. 21, no 1, , p. 35-49 (ISSN 1573-6873, PMID 16633938, DOI 10.1007/s10827-006-7074-5, S2CID 8911457, lire en ligne)
- Kostal L, Lansky P, Rospars JP, « Neuronal coding and spiking randomness », Eur. J. Neurosci., vol. 26, no 10, , p. 2693-701 (PMID 18001270, DOI 10.1111/j.1460-9568 .2007.05880.x, S2CID 15367988)
- Nitin Gupta, Swikriti Saran Singh et Mark Stopfer, « Fenêtres d'intégration oscillatoire dans les neurones », Nature Communications, vol. 7, , p. 13808 (ISSN 2041-1723, PMID 27976720, PMCID 5171764, DOI 10. 1038/ncomms13808, Bibcode 2016NatCo...713808G)
- E. Geoffrois, J.M. Edeline et J.F. Vibert, Computation in Neurons and Neural Systems, Springer, , 133-8 p. (ISBN 978-0-7923-9465-5), « Learning by Delay Modifications ».
- Sjöström, Jesper, et Wulfram Gerstner. " Spike-timing dependent plasticity ". Spike-timing dependent plasticity 35 (2010).
- T. Gollisch et Meister, M., « Rapid Neural Coding in the Retina with Relative Spike Latencies », Science, vol. 319, no 5866, , p. 1108-1111 (PMID 18292344, DOI 10.1126/science.1149639, Bibcode 2008Sci...319.1108G, S2CID 1032537, lire en ligne)
- Gilles Wainrib, Michèle, Thieullen et Khashayar, Pakdaman, « Variabilité intrinsèque de la latence au premier pic », Biological Cybernetics, vol. 103, no 1, , p. 43-56 (PMID 20372920, DOI 10. 1007/s00422-010-0384-8, S2CID 7121609)
- Johnathan D Victor, « Spike train metrics », Current Opinion in Neurobiology, vol. 15, no 5, , p. 585-592 (PMID 16140522, PMCID 2713191, DOI 10.1016/j.conb.2005.08.002)
- Robert M. Hallock et Patricia M. Di Lorenzo, « Codage temporel dans le système gustatif », Neuroscience & Biobehavioral Reviews, vol. 30, no 8, , p. 1145-1160 (PMID 16979239, DOI 10.1016/j.neubiorev.2006 .07.005, S2CID 14739301)
- Alan Carleton, Riccardo Accolla et Sidney A. Simon, « Coding in the mammalian gustatory system », Trends in Neurosciences, vol. 33, no 7, , p. 326-334 (PMID 20493563, PMCID 2902637, DOI 10.1016/j.tins.2010 .04.002)
- Tregub, S. (2021). "Algorithme de l'esprit : Théorie de la transduction téléologique." in Symphonie de la matière et de l'esprit. (ISBN 9785604473948)
- Rose, S.P.R. (14 février 2015) 50 ans de neurosciences. Lancet, 385(9968), 598-9. https://doi.org/10.1016/S0140-6736(15)60224-0
- Trettenbrein PC (2016) The demise of the synapse as the locus of memory: a looming paradigm shift?[La disparition de la synapse en tant que lieu de la mémoire : un changement de paradigme imminent ?] Frontiers in Systems Neuroscience, 10: 88. https://doi.org/10.3389/fnsys.2016.00088
- Traill, R.R. (2008/2005b). Penser par Molécule, Synapse ou toutes les deux ? — Du schéma de Piaget, à la Sélection/Rédaction du ARNnc. Gen.Sci.J., https://www.gsjournal.net/Science-Journals/Research Papers/View/892
- Traill R.R. (2022) Coding for the Brain: RNA, its Photons, and Piagetian Higher-Intelligence through Action. Journal of Psychiatry and Psychiatric Disorders, 6, 276-297. https://doi.org/10.26502/jppd.2572-519X0175
- Traill R.R. (2022) Codage pour le cerveau : l'ARN, ses photons et l'intelligence supérieure piagétienne par l'action. https://www.ondwelle.com/CodageCerveau(Fr).pdf
- Traill, R.R. (1988). The case that mammalian intelligence is based on sub-molecular memory coding and fibre-optic capabilities of myelinated nerve axons. Speculations in Science and Technology, 11(3), 173-181. https://www.ondwelle.com/OSM10en.pdf
- Mattick, J.S. (2001). Les ARN non codants : les architectes de la complexité eucaryote. Rapports EMBO, 2(11), 986-991. http://emboreports.npgjournals.com/cgi/content/full/2/11/986
- Sun Y[an], Chao Wang, & Jiapei Dai (janvier 2010). "Les bio-photons en tant que signaux de communication neuronaux démontrés par l'autographie de biophotons in situ". Photochem. Photobiol. Sci., 9, 315-322. https://doi.org/10.1039/b9pp00125e
- Zangari A., D.Micheli, R.Galeazzi & A.Tozzi, V.Balzano, G.Bellavia & M.E.Caristo (2021) "Photons détectés dans le nerf actif par technique photographique" Scientific Reports, 11, 3022. https://doi.org/10.1038/s41598-021-82622-5
- Callahan, PS (1977) "Tuning in to Nature". Routledge & Kegan Paul : Londres.
- Traill, R.R. (2005c). How Popperian positivism killed a good-but-poorly-presented theory — Insect Communication by Infrared. Gen.Sci.J., https://www.gsjournal.net/Science-Journals/Research%20Papers/View/897
Lectures complémentaires
- Földiák P, D Endres, codage Sparse, Scholarpedia, 3 (1): 2984, 2008.
- Dayan P & Abbott LF. Theoretical Neuroscience: Computational and Mathematical Modeling of Neural Systems. Cambridge, Massachusetts: The MIT Press; 2001. (ISBN 0-262-04199-5)
- Rieke F, D Warland, Ruyter van Steveninck R, Bialek W. Spikes: Exploring the Neural Code. Cambridge, Massachusetts: The MIT Press; 1999. (ISBN 0-262-68108-0)
- Olshausen et Field, « Emergence of simple-cell receptive field properties by learning a sparse code for natural images », Nature, vol. 381, no 6583, , p. 607–9 (PMID 8637596, DOI 10.1038/381607a0, Bibcode 1996Natur.381..607O)
- Tsien, « On initial Brain Activity Mapping of episodic and semantic memory code in the hippocampus », Neurobiology of Learning and Memory, vol. 105, , p. 200–210 (PMID 23838072, PMCID 3769419, DOI 10.1016/j.nlm.2013.06.019)