Cellule pariétale
Les cellules pariétales (ou oxyntiques ou cellules bordantes) font partie des différentes cellules composant la paroi des glandes gastriques. Elles sécrètent de l'acide chlorhydrique (HCl) grâce aux pompes H+/K+ ATPase et aux canaux chlore, ainsi que le facteur intrinsèque, nécessaire à l'absorption de la vitamine B12.
Localisation
Les cellules pariétales sont présentes dans les glandes fundiques de la muqueuse gastrique. Ces glandes fundiques sont des glandes exocrines multicellulaires (plusieurs types cellulaires différents) dont le produit de sécrétion est destiné à la surface interne de l'estomac. Les cellules pariétales sont présentes dans le collet et la partie supérieure de ces glandes fundiques et exhibent une morphologie assez particulière qui leur prévaut parfois le nom de "cellule à œuf sur le plat".
Description
Les cellules pariétales sont généralement assez larges, d'apparence arrondie à triangulaire, avec un noyau central et sphérique. Elles ont une position assez excentrée par rapport aux cellules. La chromatine est assez condensée, le cytoplasme est éosinophile et l'appareil de protéosynthèse (Golgi, et RER) sont moyennement développés. Les cellules présentent également de nombreuses mitochondries, en relation avec leur fonction de sécrétion d'acide chlorhydrique via la pompe à protons.
Ces cellules pariétales présentent plusieurs caractéristiques ultrastructurales qui les démarquent fortement des autres types cellulaires des glandes fundiques :
- Un système canaliculaire intracellulaire : la membrane plasmique apicale s'invagine pour former des canalicules ("petits canaux") tubulaires dont la surface est considérablement augmentée par la présence de plusieurs milliers de microvillosités. L'extensivité et le nombre de ces microvillosités reflète par ailleurs l'activité métabolique de ces cellules
- Un système tubulo-vésiculaire : le cytoplasme contient de nombreuses vésicules qui renferment les pompes à protons (pompes H+/K+/ATPases) qui viennent fusionner avec la membrane des canalicules lors d'une stimulation. Ces vésicules intracytoplasmiques sont généralement situées dans le cytoplasme qui est immédiatement adjacent aux canalicules. Lors d'une stimulation (e.g, arrivée du bol alimentaire dans l'estomac et distension de sa paroi), ces vésicules fusionnent rapidement avec la membrane des canalicules, et des microvillosités apparaissent. On observe donc une augmentation considérable de la surface, et de l'activité de sécrétion d'acide, ainsi qu'une diminution du nombre de tubulo-vésicules dans le cytoplasme à la suite d'une stimulation.
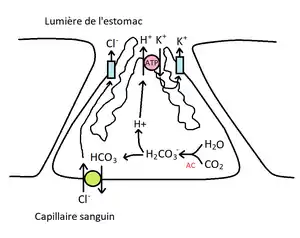
Au niveau du pôle (côté) apical (luminal) de la cellule, il y a des canaux potassium, des canaux chlore et des pompes H+/K+/ATPase. Au niveau basolatéral il y a une pompe HCO3-/Cl- et on y trouve des récepteurs comme le récepteur histaminergique de type 2 ou H2 qui est un RCPG couplé à une proteine Gs, un récepteur muscarinique de type 3 ou M3 (RCPG) métabotropique couplé avec Gq/11 qui active une phospholipase C (effecteur primaire), un récepteur de la gastrine CCK-8 qui active aussi une phospholipase C, un récepteur de la somatostatine couplé à une protéine Gi (inhibitrice), un récepteur de la prostaglandine PGE-2 couplé à une protéine Gi et un récepteur de l'EGF et du TGFα qui est couplé aussi à une protéine Gi qui diminue la synthèse d'AMPc.
Activité
La pompe à protons (H+/K+/ATPase) est une pompe présente au niveau de la membrane des canalicules. Elle permet la sécrétion active de protons H+, contre la rentrée de K+ et la dépense d'un ATP. Les cellules pariétales peuvent sécreter le H+ contre un gradient électrochimique 10^6 fois supérieur dans la lumière. Ce gradient est maintenu par la présence de jonctions serrées qui empêchent le H+ de traverser la muqueuse en voie paracellulaire et réintégrer la circulaire sanguine. La cellule puise ses protons (H+) grâce à l'action d'une enzyme, l'anhydrase carbonique, qui permet de former du H+ et HCO3- à partir de CO2 et d'eau.
H2O + CO2 ↔ H2CO3 ↔ HCO3− + H+.
Le H+ ainsi formé peut ainsi être expulsé au pôle apical par la pompe à proton, où il intègre la lumière des glandes fundiques pour se retrouver finalement dans la lumière de l'estomac et vient se mélanger au bol alimentaire. Le HCO3- formé est expulsé au pôle basal de la cellule grâce à l'échangeur HCO3-/Cl-. Le Cl- passe au pôle apical et traverse la membrane apicale pour se joindre aux produits de sécrétions via une conductance au chlore. Le produit de sécrétion des cellules pariétales est donc le HCl, bien qu'il soit directement dissocié en H+ et Cl-.
Le HCl est un acide fort : sa dissociation est considérée comme complète. On pourrait donc se demander ce que vient faire le Cl- comme produit de sécrétion dans ce cas, puisqu'il ne participe pas à l'acidité de l'estomac en tant que tel : il joue un rôle d'électroneutralité. En sécrétant des H+ et des Cl-, à stoechiométrie égale, on a une électroneutralité. Dès lors, la pompe à proton ne doit faire face qu'au gradient de concentration augmentant de H+, et non pas au gradient électrique qui pourrait augmenter s'il n'y avait pas une telle électroneutralité, et le cout énergétique associé serait bien plus élevé.
De manière à maintenir la pompe à proton active, le K+ pompé dans la cellule peut refuire par une simple conductance à K+ au pôle apical.
La fusion des vésicules avec la membrane plasmique est dépendante de différents stimuli, et donc l'activité sécrétoire de la cellule pariétale est dite régulée. Cette régulation se fait par le niveau intracellulaire de cAMP, un messager secondaire important produit grâce à l'adénylate cyclase. Cette protéine membranaire - qui réside dans le feuillet interne - est modulée par différentes voies de signalisations. Au pôle basal, la cellule présente plusieurs récepteurs qui peuvent moduler l'activité sécrétrice des cellules pariétales en modifiant l'activité de l'adénylate cyclase.
Plusieurs facteurs stimulent la sécrétion d'acide :
- Nerveux : des stimulations parasympathiques, cheminées par le nerf vague (NC X), agissent directement sur les cellules pariétales via leur neurones postganglionnaires situés dans la paroi et associés au plexus de Meissner (ganglions intrapariétaux). La libération d'acétylcholine par ces fibres efférentes parasympathiques entraine l'activation des cellules pariétales. La stimulation peut également se faire par voie indirecte, en stimulant les cellules entéroendocrines sécrétant la gastrine (estomac et duodénum), et l'histamine via les cellules ECL, ou encore l'inhibition de la libération de somatostatine. L'acétylcholine agit par voie directe en se liant à des récepteurs muscarinique M3 (Récepteur Couplé à une Protéine G, RCPG).
- Hormonal : plusieurs hormones ou agents paracrines peuvent influencer la sécrétion d'acide. La gastrine est un agent très important qui stimule activement la production d'acide. L'insuline joue également un rôle important.
- Histamine : l'histamine est sécrété par les cellules ECL et agit sur les cellules pariétales. Elle agit en se liant à des récepteurs histaminergiques H2, des RCPGs. La protéine G, une Gs (pour G stimulatrice) - lorsqu'elle est activée par liaison de l'histamine à son récepteur - stimule la production d'AMPc par l'adénylate cyclase.
- La caféine agit sur la sécrétion acide en agissant comme un inhibiteur de la phosphodiestérase, augmentant ainsi le cAMP dans les cellules pariétales.
Références