Test de foraminifères
Les tests de foraminifères sont les tests (ou coquilles) de foraminifères.
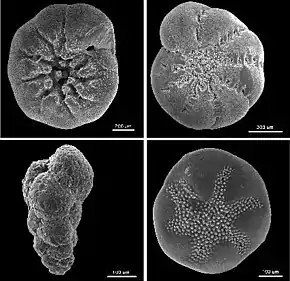
Les foraminifères (Foraminifera en latin) sont des organismes eucaryotes unicellulaires prédateurs, principalement marins, et généralement protégés par une coquille. Celle-ci peut être constituée d'une seule chambre ou avoir de multiples cavités interconnectées. La machinerie cellulaire est contenue dans cette coquille. Le test est tellement important à la biologie du groupe taxonomique qu'il lui donne son nom scientifique — foraminifera, du latin "foramen" trou, "qui porte des trous" - en référence aux pores connectant les chambres de la coquille de certaines espèces.
Test est un mot venant du latin testa, « récipient arrondi », qui a donné aussi le terme « tête ».
Les tests de foraminifères sont généralement composés de calcite, une forme de carbonate de calcium (CaCO3), mais peuvent être faits parfois d'aragonite, de l'agglutination de particules de sédiment, de chitine ou, plus rarement, de silice[1]. D'autres foraminifères sont totalement dépourvus de tests[2].
La dureté de la plupart des tests de foraminifères conduit à une excellente conservation à l'état fossile. Les tests sont donc utilisés en recherche comme sources d'information au sujet du climat du passé (Paléoclimatologie) et des environnements disparus[3], mais aussi pour la taxonomie du groupe.
Biologie
Il y a plus de 50 000 espèces reconnues, que ce soit des espèces actuelles (entre 6 700 et 10 000)[4] - [5] ou des espèces fossiles (environ 40 000)[6] - [7].
Elles mesurent généralement moins de 1 mm, mais certaines espèces sont bien plus grandes. L'espèce la plus grande fait partie de la super-famille des xénophyophores et atteint la taille de 20 cm[8].
La plupart des foraminifères sont benthiques, mais environ 40 espèces actuelles sont planctoniques[9].
Le test sert à la protection de l'organisme qui vit à l'intérieur.
Les ouvertures dans le test qui permettent au cytoplasme de s'étendre à l'extérieur sont appelées apertures[10]. L'aperture primaire, conduisant à l'extérieur, peut prendre des formes variées chez les différentes espèces: ronde, en croissant, en fente, radiale (en étoile), dendritique (avec des branchements). Certains foraminifères ont des apertures primaires "dentées", ou présentant des lèvres. Il peut n'y avoir qu'une seule aperture primaire ou de multiples. Dans ce deuxième cas, elles peuvent être groupées ou équatoriales. En plus des apertures primaires, certaines espèces ont des apertures supplémentaires. Elles peuvent être présentes sous forme relictuelle (une aperture primaire d'une étape précédente de développement) ou être des structures uniques.
La forme du test est très variable parmi les différents foraminifères. Ceux-ci peuvent présenter une seule chambre (uniloculaire) ou de multiples chambres (multiloculaire). Dans les formes multiloculaires, les nouvelles chambres s'ajoutent lorsque l'organisme grandit. Une vaste variété de morphologies des tests est trouvée que ce soit chez les formes uniloculaires ou multiloculaires, incluant des formes spiralées, sériées, et celle des Miliolina, entre autres[11].
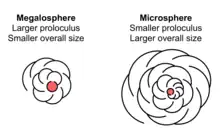
De nombreux foraminifères montrent un dimorphisme dans leurs tests, avec des individus soit mégalosphérique soit microsphérique. Ces noms fontt référence à la taille de la chambre initiale (ou proloculus), plutôt qu'à la taille de l'organisme entier.
Les tests, en tant que fossiles, sont connus en paléontologie, les plus anciens datant de la période de l'Édiacarien[12], et de nombreux sédiments marins en sont composés principalement. Par exemple, le calcaire des pyramides d'Égypte est composé presque entièrement de foraminifères benthiques nummulitiques[13]. Il est estimé que les foraminifères des récifs générent environ 43 million de tonnes de carbonate de calcium par an[14].
Des études génétiques ont mis en évidence que le genre Reticulomyxa et les espèces de la super-famille des xénophyophores sont des foraminifères sans tests. Quelques autres espèces, ressemblant à des amibes produisant des pseudopodes réticulés, ont été auparavant classées dans le groupe des Granuloreticulosa, mais celui-ci n'est plus considéré comme un groupe naturel, et la plupart des espèces sont maintenant placées parmi les Cercozoa[15].
Composition
La forme et la composition des tests est le moyen principal d'identifier et de classer les foraminifères. La plupart secrète des tests calcaires, composés de carbonate de calcium[16]. Les tests calcaires peuvent être composés soit d'aragonite soit de calcite selon les espèces. Parmi celles à test de calcite, le test peut contenir une substitution plus ou moins importante par du magnésium[17]. Le test contient une matrice organique, qui peut parfois être recouvrée d'échantillons fossiles[17].
Des études suggèrent l'existence d'un certain niveau d'homoplasie chez les foraminifères, et que, ni les espèces à tests agglutinés, ni celles à tests calcaires, ne forment des groupes monophylétiques[18].
Test mou
Chez certaines espèces, les tests peuvent être composés de matériau organique, constitué typiquement de la protéine tectine. Les parois à tectine peuvent contenir des particules de sédiment adhérant de façon lâche à la surface[11].
Les foraminifères du genre Reticulomyxa n'ont pas du tout de test, ayant une paroi cellulaire membraneuse[2]. Les foraminifères à paroi organique ont traditionnellement été groupés parmi les "Allogromiidae". Cependant, les études génétiques ont montré que ce n'est pas un groupe naturel[18].
Test agglutiné
D'autres foraminifères ont des tests faits de petits morceaux de sédiment cimentés ensemble (agglutinés) soit par des protéines (possiblement ressemblant au collagène), du carbonate de calcium , ou de l'oxyde de fer (III)[11] - [19]. Dans le passé, ces formes été regroupées pour les spécimens à une seule chambre en tant qu' "Astrorhizidae" et pour ceux à chambres multiples en tant que Textulariida. Cependant, des études génétiques récentes suggèrent que les "astrorhizidés" ne forment pas un groupe naturel, mais forment plutôt une large base à l'arbre phylogénétique des foraminifères[18].
Les foraminifères du groupe des Textulariida, à la différence des autres membres actuels des Globothalamea, ont des tests agglutinés. Cependant, les grains de ces tests sont cimentés avec de la calcite. Ces tests contiennent de petits nanoparticules (<100 nm) globulaires, similaires à celles chez les autres Globothalamea. Ces tests peuvent aussi avoir de nombreux pores, autre caractéristique les unifiant aux Globothalamea[20].
Les foraminifères à tests agglutinés peuvent être sélectifs quant au choix des particules qu'ils incorporent dans leurs coquilles. Certaines espèces préfèrent certaines tailles et types de particules rocheuses; d'autres espèces ont des préférences pour certains matériaux biologiques. Certaines espèces agglutinent préférentiellement des coccolithes pour former leurs tests; d'autres utilisent des plaques d'échinodermes, des diatomées, ou même d'autres tests de foraminifères[21]. L'espèce Spiculosiphon oceana agglutine préférentiellement des spicules de silice d'éponges en utilisant un ciment organique. Elle montre aussi une grande sélectivité à l'encontre de la forme des spicules, utilisant ceux allongés pour son "pied" et de plus courts pour son "bulbe". L'hypothèse a été émise qu'elle utilise les spicules à la fois comme un moyen de s'élever du fond de la mer et pour allonger la portée de ses pseudopodes pour capturer des proies[19].

Les tests agglutinés de xénophyophores sont les plus grands de tous les foraminifères, atteignant la taille de 20 cm de diamètre. Le nom "xénophyophore", signifiant "porteur de corps étrangers", fait référence à cette habitude d'agglutination. Les xénophyophores utilisent sélectivement des grains de sédiment d'une taille située entre 63 et 500 µm, en évitant les galets plus grands et les boues plus fines. Le type de sédiment semble être un facteur important dans la façon dont les particules sont agglutinées. Ces types de particules incluent préférentiellement des sulfites, des oxydes, des verres volcaniques et des tests de foraminifères plus petits. Des xénophyophores de 1,5 cm de diamètre ont été trouvés complètement nus, sans aucune forme de test[22].
Test calcaire
Certains foraminifères produisent des tests calcaires. Plusieurs structures de cristaux de calcite différents existent.
Test porcelané

Les parois des tests porcelané sont trouvés dans le groupe des Miliolida. Elles sont formées d'une calcite à haute teneur en magnésium organisée avec deux faces ordonnées, extérieure et intérieure (l extrados et l intrados, respectivement), et des cristaux de calcite en forme d'aiguilles orientés aléatoirement, formant une couche épaisse au centre (la "porcelaine"). Une couche interne organique est égalemnt présente. La surface externe peut présenter une structure en creux, mais elle n'est pas perforée de trous. Les Miliolidae du groupe des "Cornuspiridae" sont marqués apparemment par l'absence d'extrados[23] - [24] - [20].
Test monocristallin
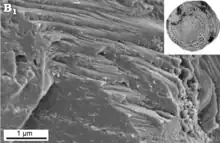
Un test "monocristallin" possède une structure traditionnellement décrite chez les Spirillinida. Cependant, ces tests restent peu connus. Certains Spirillinidae supposés "monocristallin" ont en fait des tests consistant en une mosaïque de très petits cristaux lorsqu'ils sont observés au microscope électronique. Les observations au microscope de Patellina sp. suggèrent cependant que le type de test monocristallin existe réellement, avec des faces de clivage apparentes[20].
Faisceau de fibres

Les tests des membres du groupe des Lagenida consistent en des « faisceaux de fibres » pouvant atteindre plusieurs dizaines de micromètres de longueur. Chaque "faisceau" est formé d'un cristal de calcite singulier. Il est de section triangulaire et possède un pore au centre (ce qu'on suppose être un artéfact du dépôt du test). There is also an internal organic layer, attached to the "cone" structure of the fibre bundles. As the crystalline structure varies significantly from that of autres foraminifères à test calcaire, it is thought to represent a évolution séparée de test calcaire. Le processus exact de la minéralisation des Lagenida demeure unclear[24].
Test hyalin
Les tests des membres du groupe des Rotaliida sont décrits comme "hyalins". Ils sont formés de "nanograins" de calcite, à teneur basse à élevée en magnésium, positionés avec leur axes perpendiculaires à la surface externe du test. De plus, ces nanograins peuvent présenter des structures de plus haut niveau, telles que des rangées, des colonnes, ou des faisceaux[20]. La paroi du test est de façon caractéristique bilamellaire (à deux couches) et perforée de petits pores. Prise en sandwich entre les deux lamelles, on trouve la couche médiane, faite de protéines. La couche médiane est très variable selon l'espèce. Elle peut être très bien délimitée ou pas. Certains genres peuvent contenir des particules de sédiment au cœur de la couche médiane[11] - [25] - [24].
Les membres du groupe éteint des Fusulinida ont traditionnellement été considérés uniques du fait de posséder des tests formés de cristaux homogénes microgranulaires sans orientation préférentielle et presque pas de ciment. Cependant, une étude de 2017 montre que la structure microgranulaire supposée est en fait le résultat d'une altération diagénétique des fossiles, et que les tests non altérés de Fusulinidae ont une structure hyaline. Cela suggère que le groupe est affilié avec les Globothalamea[26].

Les membres du groupe des Robertinida ont des tests formés d'aragonite avec des perforations. Ils sont similaires aux tests des Rotaliidae dans le sens qu'ils sont formés de nanograins. Cependant, ils différent en composition et avec le fait qu'ils possèdent des domaines columnaires bien organisés. Du fait que les premiers foraminifères planctoniques avaient des tests d'aragonite, il a été suggéré que cela soit une évolution separée d'un style de vie planctonique au sein des Robertinida, plutôt que d'être des parents des Globigerinana[20].
Les tests hyalins d'aragonite sont également présents chez les Involutinida[24].
Test à spicules

Les Carterinida, incluant les genres Carterina et Zaninettia, ont une structure cristalline unique du test ce qui a compliqué depuis longtemps leur classification. Le test consiste en spicules de calcite à basse teneur de magnésium, liés à une matrice organique et contenant des morceaux de matière organique. Ceci a conduit certains chercheurs à conclure que le test est du type agglutiné. Cependant, les études en milieu naturel n'ont pas pu mettre en évidence cette agglutination, et, en fait, les représentants des genres ont été découverts sur un substrat artificiel où les particules de sédiment ne s'accumulent pas[27]. Une étude génétique de 2014 a montré que les carterinida sont une lignée indépendante au sein des Globothalamea, et conforte l'idée que les spicules secrétés ont des formes différentes entre les spécimens de Carterina et Zaninettia collectés au niveau de la même localité (ovoïdes chez Carterina, ronds-rectangulaires chez Zaninettia)[28].
Silicate
Un genre, Miliamellus, a un test non perforé composé de silice opalescente[29]. Il est similaire en forme et structure aux tests porcelanés des Miliolidae typiques. Le test consiste en une couche organique interne et externe, et en une couche médiane de silice formés de batonnets allongés. La couche de silice est encore divisée entre sous-unités internes, médianes et externes. Les sous-unités externes et internes sont approximatement de 0,2 μm d'épaisseur et consistent en feuillets sub-parallèles de batonnets de silice avec leur axe long parallèle à la surface du test. La sous-unité médiane fait approximatement 18 μm d'épaisseur et consiste en une structure tridimensionelle de batonnets de silice sans composante organique dans l'espace ouvert. L'ultrastructure diffère de celle des Miliolida dans le sens que les batonnets sont deux fois plus longs et deux fois plus épais en moyenne, et que les batonnets de Miliamellus sont creux plutôt que pleins, et, évidemment, ont un test de silice plutôt que de calcite[30].
Construction de la paroi du test
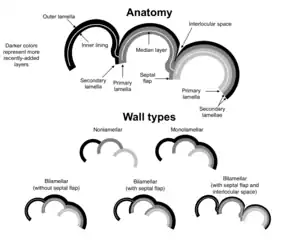
Lorsqu'un test secrété est présent, les parois peuvent être non lamellaire ou lamellaire.
Les parois non lamellaires sont trouvées chez certains forminifères, tels que les Carterinida, les Spirillinida et les Miliolida. Chez ces formes, la secrétion d'une nouvelle chambre n'est pas associée avec un dépôt sur une chambre précédente. En ce sens, il n'y a pas de dépôt associé de couches de calcite sur le test[25].
Chez les foraminifères à parois lamellaires, le dépôt pour une nouvelle chambre est accompagnée du dépôt d'une couche au-dessus des chambres précédemment formées. Cette couche peut recouvrir toutes les chambres précédentes, ou seulement certaines d'entre elles. Ces couches sont appelées lamelles secondaires.
Les foraminifères à parois lamellaires peuvent être encore divisés entre ceux avec des parois monolamellaires et ceux avec des parois bilamellaires. Les foraminifères monolamellaires secrétent des parois qui consistent en une couche unique, tandis que ceux du type bilamellaire ont une double couche avec une couche médiane organique, contenant parfois des particules de sédiment. Les foraminifères monolamellaires incluent les Lagenida, tandis que les bilamellaires incluent les Rotaliida (incluant les sous-groupes planctoniques majeurs, les Globigerinina)[25].
Les parois des tests bilamellaires peuvent être encore divisées entre celles avec des volets septaux (une couche de la paroi du test recouvrant le septum précédemment secrété) et celles n'ayant pas de volets septaux. Les volets septaux sont connus chez tous les foraminifères autres que ceux avec des parois bilamellaires.
La présence d'un volet septal est souvent, bien que pas toujours, assocé à la présence d'un espace interloculaire. Comme le nom le suggère, c'est un petit espace localisé entre les chambres. Celui-ci peut être ouvert et faire partie de la surface externe du test, ou il peut être fermé pour former un vide. La couche autour de ce vide est formée de différentes parties des lamelles dans différents genres, suggérant une évolution indépendante des espaces interloculaires pour réenforcer le test[25].
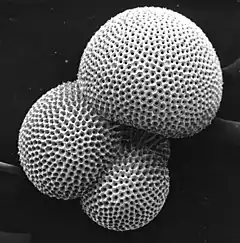


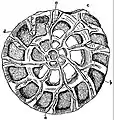 Section montrant les chambres d'un foraminifère en spirale (Rotalia beccarii).
Section montrant les chambres d'un foraminifère en spirale (Rotalia beccarii). Individu vivant d' Ammonia tepida émettant un ectoplasme granulaire pour attraper de la nourriture.
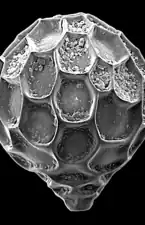
Individu vivant d' Ammonia tepida émettant un ectoplasme granulaire pour attraper de la nourriture. Groupe de foraminifères planctoniques du Golfe du Mexique.
Groupe de foraminifères planctoniques du Golfe du Mexique. Test de Baculogypsina sphaerulata, constituant du sable étoilé.
Test de Baculogypsina sphaerulata, constituant du sable étoilé. Fossiles de Nummulitidae de tailles variées datant de l'Éocène.
Fossiles de Nummulitidae de tailles variées datant de l'Éocène.
Références
- (en) J.P. Kennett et M.S. Srinivasan, Neogene planktonic foraminifera: a phylogenetic atlas, Hutchinson Ross, (ISBN 978-0-87933-070-5, lire en ligne)
- (en) Jan Pawlowski, Ignacio Bolivar, Jose F. Fahrni, Colomban De Vargas et Samuel S. Bowser, « Molecular Evidence That Reticulomyxa Filosa Is A Freshwater Naked Foraminifer », Journal of Eukaryotic Microbiology, vol. 46, no 6, , p. 612–617 (ISSN 1550-7408, PMID 10568034, DOI 10.1111/j.1550-7408.1999.tb05137.x, S2CID 36497475)
- Wassilieff Maggy, 2006. "Plankton - Animal plankton", Te Ara - the Encyclopedia of New Zealand. Accessed: 2 November 2019.
- (en) J. Pawlowski, F. Lejzerowicz et P. Esling, « Next-Generation Environmental Diversity Surveys of Foraminifera: Preparing the Future », The Biological Bulletin, vol. 227, no 2, , p. 93–106 (ISSN 0006-3185, PMID 25411369, DOI 10.1086/BBLv227n2p93, S2CID 24388876, lire en ligne)
- (en) Adl, S.M. et al., 2007. Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI 10.1080/10635150701494127.
- (en) Pawlowski, J., Lejzerowicz, F., & Esling P., 2014. Next-generation environmental diversity surveys of foraminifera: preparing the future. The Biological Bulletin, 227(2), 93-106.
- « World Foraminifera Database »
- Marshall M., « Zoologger: 'Living beach ball' is giant single cell », New Scientist, (lire en ligne)
- (en) C. Hemleben, O.R. Anderson et M. Spindler, Modern Planktonic Foraminifera, Springer-Verlag, (ISBN 978-3-540-96815-3, lire en ligne)
- (en) C Lana, « Cretaceous Carterina (Foraminifera) », Marine Micropaleontology, vol. 41, nos 1–2, , p. 97–102 (DOI 10.1016/S0377-8398(00)00050-5, Bibcode 2001MarMP..41...97L)
- (en) Pratul Kumar Saraswati et M. S. Srinivasan, Calcareous-Walled Microfossils, Springer International Publishing, , 81–119 p. (ISBN 978-3-319-14574-7, DOI 10.1007/978-3-319-14574-7_6)
- (en) A. E. Kontorovich, A. I. Varlamov, D. V. Grazhdankin, G. A. Karlova, A. G. Klets, V. A. Kontorovich, S. V. Saraev, A. A. Terleev, S. Yu. Belyaev, I. V. Varaksina et A. S. Efimov, « A section of Vendian in the east of West Siberian Plate (based on data from the Borehole Vostok 3) », Russian Geology and Geophysics, vol. 49, no 12, , p. 932–939 (ISSN 1068-7971, DOI 10.1016/j.rgg.2008.06.012, Bibcode 2008RuGG...49..932K, lire en ligne)
- (en) Foraminifera: History of Study, University College London, site visité le 20 septembre 2007
- (en) M. R. Langer, Silk, M. T. B. et Lipps, J. H., « Global ocean carbonate and carbon dioxide production: The role of reef Foraminifera », Journal of Foraminiferal Research, vol. 27, no 4, , p. 271–277 (DOI 10.2113/gsjfr.27.4.271, lire en ligne)
- (en) Adl S. M., Simpson, A. G. B., Farmer, M. A., Anderson et al., « The new higher level classification of Eukaryotes with emphasis on the taxonomy of Protists », Journal of Eukaryotic Microbiology, vol. 52, no 5, , p. 399–451 (PMID 16248873, DOI 10.1111/j.1550-7408.2005.00053.x
 , S2CID 8060916)
, S2CID 8060916) - (en) Barun K. Sen Gupta, Foraminifera: notes for a short course organized by M.A. Buzas and B.K. Sen Gupta, vol. 6, University of Tennessee, Dept. of Geological Sciences, coll. « Studies in Geology », , 37–50 p. (ISBN 978-0910249058, OCLC 9276403), « Ecology of benthic Foraminifera »
- (en) Barun K. Sen Gupta, Modern Foraminifera, Springer Netherlands, , 7–36 p. (ISBN 978-0-306-48104-8, DOI 10.1007/0-306-48104-9_2), « Systematics of moder Foraminifera »
- (en) Jan Pawlowski, Maria Holzmann et Jarosław Tyszka, « New supraordinal classification of Foraminifera: Molecules meet morphology », Marine Micropaleontology, vol. 100, , p. 1–10 (ISSN 0377-8398, DOI 10.1016/j.marmicro.2013.04.002, Bibcode 2013MarMP.100....1P, lire en ligne)
- (en) Manuel Maldonado, María López-Acosta, Cèlia Sitjà, Ricardo Aguilar, Silvia García et Jean Vacelet, « A giant foraminifer that converges to the feeding strategy of carnivorous sponges: Spiculosiphon oceana sp. nov. (Foraminifera, Astrorhizida) », Zootaxa, vol. 3669, no 4, , p. 571–584 (ISSN 1175-5334, PMID 26312358, DOI 10.11646/zootaxa.3669.4.9, hdl 10261/92975 , lire en ligne)
- (en) Zofia Dubicka, « Chamber arrangement versus wall structure in the high-rank phylogenetic classification of Foraminifera », Acta Palaeontologica Polonica, vol. 64, (ISSN 0567-7920, DOI 10.4202/app.00564.2018 )
- (en) Erik Thomsen et Tine L. Rasmussen, « Coccolith-Agglutinating Foraminifera from the Early Cretaceous and How They Constructed Their Tests », Journal of Foraminiferal Research, vol. 38, no 3, , p. 193–214 (ISSN 0096-1191, DOI 10.2113/gsjfr.38.3.193, lire en ligne)
- (en) Lisa A. Levin et Cynthia L. Thomas, « The ecology of xenophyophores (Protista) on eastern Pacific seamounts », Deep Sea Research Part A. Oceanographic Research Papers, vol. 35, no 12, , p. 2003–2027 (ISSN 0198-0149, DOI 10.1016/0198-0149(88)90122-7, Bibcode 1988DSRA...35.2003L)
- (en) Sreepat Jain, Benthic Foraminifera, New Delhi, Springer India, coll. « Springer Geology / Fundamentals of Invertebrate Palaeontology: Microfossils », , 171–192 p. (ISBN 978-81-322-3962-8, DOI 10.1007/978-81-322-3962-8_9)
- (en) Zofia Dubicka, Krzysztof Owocki et Michał Gloc, « Micro- and Nanostructures of Calcareous Foraminiferal Tests: Insight from Representatives of Miliolida, Rotaliida and Lagenida », Journal of Foraminiferal Research, vol. 48, no 2, , p. 142–155 (ISSN 0096-1191, DOI 10.2113/gsjfr.48.2.142, lire en ligne)
- (en) Hans Jørgen Hansen, Shell construction in modern calcareous Foraminifera, Dordrecht, Springer Netherlands, , 57–70 p. (ISBN 978-0-306-48104-8, DOI 10.1007/0-306-48104-9_4)
- (en) Zofia Dubicka et Przemysław Gorzelak, « Unlocking the biomineralization style and affinity of Paleozoic fusulinid foraminifera », Scientific Reports, vol. 7, no 1, , p. 15218 (ISSN 2045-2322, PMID 29123221, PMCID 5680253, DOI 10.1038/s41598-017-15666-1, Bibcode 2017NatSR...715218D)
- (en) Altair Machado et Facelucia Barros, « The occurrence of Carterina spiculotesta (Carter, 1877) on an artificial substrate », Check List, vol. 9, no 4, , p. 813–814 (ISSN 1809-127X, DOI 10.15560/9.4.813 , lire en ligne)
- (en) Jan Pawlowski, Maria Holzmann et Jean-Pierre Debenay, « Molecular Phylogeny of Carterina Spiculotesta and Related Species from New Caledonia », Journal of Foraminiferal Research, vol. 44, no 4, , p. 440–450 (ISSN 0096-1191, DOI 10.2113/gsjfr.44.4.440, lire en ligne)
- (en) Barun K. Sen Gupta, Modern Foraminifera, Springer, (ISBN 978-1-4020-0598-5, lire en ligne), p. 16
- (en) J Resig, H Lowenstam, R Echols et S Weiner, « An extant opaline foraminifer: test ultrastructure, mineralogy, and taxonomy. », Special Publications of the Cushman Foundation for Foraminiferal Research., vol. 19, , p. 205–214
- Favulina hexagona European Geosciences Union, 9 November 2020.
- Foraminifera: History of Study, University College London. Site visité le 18 novembre 2019.