Rythmes circalunaires des animaux marins
Plusieurs espèces d’animaux marins possèdent des rythmes biologiques circalunaires, c’est-à-dire, d’une période d’environ 29,5 jours, ce qui correspond à un cycle des phases de la Lune. Ces rythmes sont souvent reliés à des événements de reproduction qui nécessitent une synchronisation pour faciliter la fécondation[2]. En effet, la plupart des espèces marines avec un cycle de reproduction circalunaire font de la fécondation externe; les gamètes mâles et femelles sont relâchés dans l’environnement et ils doivent se rencontrer pour effectuer la fécondation[2]. Il est possible que ces espèces marines aient acquis ces rythmes au fil de l’évolution, car le relâchement des gamètes à un moment précis représente un avantage adaptatif : si les gamètes ne sont pas relâchés en même temps, les chances de fécondation seraient réduites[2].

Chez les coraux

Plusieurs espèces de coraux relâchent leurs gamètes la nuit une ou deux fois par année, 4 à 5 nuits après une pleine lune[4]. Chez le corail Acropora millepora, la transcription de certains gènes de l’horloge circadienne varie selon les phases de la Lune, ce qui pourrait indiquer que l’intensité lumineuse de la pleine lune peut induire les événements de relâchement de gamètes au coucher du Soleil à un temps précis[5]. Par exemple, les gènes circadiens Cry1, Cry2, Clock, Cycle/bmal, Timeless et Eva ont des patrons d’expression qui varient selon les phases de la Lune[5]. Ces variations peuvent être divisées en deux catégories. Le gène Cry1 démontre des amplitudes différentes de transcription entre la nouvelle lune et la pleine lune, alors que les gènes Cry2, Clock et Timeless gardent la même amplitude, mais les phases de la Lune induisent un changement de phase de leur cycle d’expression[5].
Le cryptochrome CRY2 d’Acropora millepora, un photorécepteur de la lumière bleue, est potentiellement une composante de l’horloge circadienne qui permettrait la synchronisation des événements de relâchement de gamètes avec la pleine lune[6]. Le gène Cry2, dont l’expression oscille de manière circadienne, démontre un niveau d’expression plus élevé à minuit lors de la pleine lune comparativement à la nouvelle lune[6].
La présence de CRY faisant partie de l’horloge circalunaire et circadienne chez les coraux permettent d’en connaître plus sur l’évolution de ces horloges chez les animaux multicellulaires, puisque les coraux sont dans un groupe phylogénétique (Anthozoaires) proche du groupe basal dans l’évolution des animaux[5].
Chez le sigan (poisson)

Le sigan à raies d'or (Siganus guttatus) est une espèce de poisson que l’on retrouve dans les milieux tropicaux. Le genre de poisson Siganus a une reproduction définie en fonction des phases lunaires. L’espèce Siganus guttatus, lui, se reproduit dans le premier quartier lunaire. Ce sont les gonades qui se développent en synchronicité à ce moment, puis elles retournent ensuite à leur état initial[8]. Le poisson repère les signaux de la Lune pour faire cette synchronisation. Des fluctuations du niveau de mélatonine en fonction de la lumière de la Lune ont été observées chez cette espèce[9]. La concentration de mélatonine dans le plasma est plus élevée pendant la nouvelle lune que pendant la pleine lune. Il est donc possible que cette hormone varie en fonction du niveau de lumière de la Lune la nuit. On pense que cette intensité lumineuse est en partie détectée par l’épiphyse, qui sert également à réguler la production de mélatonine[10]. Pour confirmer cette hypothèse, des tests ont été menés sur des épiphyses observées indépendamment du poisson[11]. Les observations montrent que la glande réagit de la même façon à la lumière de la Lune. L’épiphyse est donc l'intermédiaire entre les signaux lumineux de la Lune et la production de mélatonine. Le mécanisme de la mélatonine n’est pas encore exactement déterminé, mais il est connu que celle-ci possède des propriétés anti-gonadotrope[12]. En résumé, l’épiphyse du sigan à raies d’or repère les signaux lumineux de la Lune dans son premier quartier, ce qui fait en sorte de réduire la quantité de mélatonine dans le plasma et ainsi augmenter la production de gonades pour la reproduction.
Chez les Annélides

Chez l’Annélide marin Platynereis dumerilii, les ARN de gènes de l’horloge circadienne qui oscillent dans des noyaux situés dans le cerveau antérieur sont aussi entraînés par la lumière nocturne[14]. La période et l’intensité des rythmes locomoteurs de ce vers (gérés par l’horloge circadienne) varient selon les phases de la Lune, sans changement chez les oscillations de l’horloge circadienne endogène. En effet, les niveaux de transcription des gènes circadiens pdp1, clock, period et timeless varient selon les phases de la Lune[14]. Ces gènes pourraient donc faire partie ou être régulés par une autre horloge qui serait circalunaire. Il se pourrait donc qu’il existe une deuxième horloge endogène, qui n’est pas circadienne, mais plutôt circalunaire, qui contrôlerait entre autres la reproduction de cet organisme. En effet, les vers continuent à effectuer des événements de reproduction rythmiques (danse nuptiale) dont la période correspond à un cycle lunaire dans des conditions constantes de nouvelle lune[14]. Ceci indique la présence d’une horloge circalunaire endogène.
De nombreux facteurs influencent la reproduction de l’espèce Typosyllis prolifera, dont la phase lunaire. Dans des études menées en 1985, il fut démontré que cette espèce possède une horloge circalunaire endogène qui peut être entraînée par la lumière de la Lune et qui permet de contrôler les cycles de stolonisation qui correspond à la production de gonades[15]. Ainsi, dans des conditions favorables et dans un intervalle de jours correspondant à la saison de reproduction, l’initiation du développement sexuel du T. prolifera se fait normalement dans les jours près du quatrième quart de lune[16]. L’horloge circalunaire entraînerait, par des mécanismes méconnus, la sécrétion d’hormones spécifiques activant la stolonisation ou l’inhibant. Cela permet un certain niveau de synchronisation entre les individus dans l'initiation de la formation de leur stolon[16].
Le développement et le comportement sexuel chez l’espèce Odontosyllis enopla, tel que sa bioluminescence, sont aussi influencés par un cycle circalunaire. C’est pendant les premières nuits suivant la pleine lune qu'ont lieu des événements de rassemblement d’épitokes dans le but d’une reproduction massive[18]. Des études menées en 2018 ont permis de faire un profil des transcrits présents dans des femelles O. enopla pendant leur activité de bioluminescence et ont pu constater que les gènes responsables pour le rythme lunaire sont similaires à ceux retrouvés dans le modèle P. dumerilii, sans plus donner de détails[19].
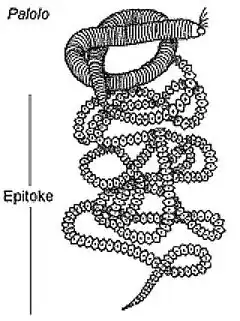
Les évènements de reproduction chez l’espèce de polychète Eunice viridis se font en accordance avec des phases lunaires précises. Il fut déterminé officiellement que c’est lors du troisième quart de lune pendant le mois de novembre ou octobre, que se détache du ver une partie de l’épitoke (le palolo) contenant les spermatozoïdes et les œufs[20]. Les détails au niveau moléculaire concernant l’émergence des palolos en relation avec les phases lunaires sont encore mal compris, des recherches doivent être faites afin de comprendre les causalités de cette périodicité[21].
Chez Clunio marinus (insecte)
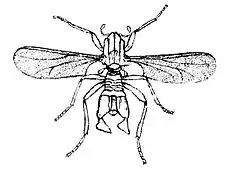
La cécidomyie intertidale (Clunio marinus) est un insecte marin qui a permis de démontrer le rôle de la Lune dans la synchronisation des rythmes de reproduction circasemilunaires[23].
Cette espèce déjoue le défi de la respiration en eau de mer en passant la majeure partie de sa vie sous forme d’œuf, puis de larve et ensuite de pupe, où ils absorbent l’oxygène par leur peau[24]. Lorsqu’il passe au stade adulte, Clunio marinus doit respirer de l’air. Les adultes ne survivent que quelques heures à marée basse, où ils doivent s’accoupler et pondre avant d’être noyés par la montée de l’eau. Les moucherons en développement émergent près des niveaux de marées basses, ce qui indique que les périodes d’accouplement et de pontes se produisent que lors des nouvelles et des pleines lunes, soit deux fois pendant le cycle lunaire, lors des marées basses[24].
Des expériences en laboratoire ont démontré que le rythme de reproduction semilunaire exprimé par la cécidomyie intertidale est un véritable rythme circasemilunaire, puisqu’il est endogène et régulé par une horloge interne[25].
L’horloge circalunaire de Clunio marinus a été étudiée par cartographie génétique, en faisant l’analyse de locus de caractères quantitatifs (LCQ ou QTL pour quantitative trait loci) sur la variation du calendrier lunaire et diurne[26]. Le gène cOps2 (ciliary opsin 2) a été identifié comme candidat potentiel impliqué dans le rythme lunaire et diurne, le gène cry1 (cryptochrome 1) dans le rythme lunaire et les gènes tim2 et tim3 (timeless) dans le rythme diurne[26].
Références
- Mariannehubert, Français : Les différentes phases de la Lune schématisées, (lire en ligne)
- (en) Kristin Tessmar-Raible, Florian Raible et Enrique Arboleda, « Another place, another timer: Marine species and the rhythms of life », BioEssays, vol. 33, no 3, , p. 165–172 (DOI 10.1002/bies.201000096, lire en ligne, consulté le )
- « Acropora millepora », sur eatlas.org.au (consulté le )
- Peter L. Harrison, Russell C. Babcock, Gordon D. Bull et James K. Oliver, « Mass Spawning in Tropical Reef Corals », Science, vol. 223, no 4641, , p. 1186–1189 (DOI 10.1126/science.223.4641.1186, lire en ligne, consulté le )
- Aisling K. Brady, Bette L. Willis, Lawrence D. Harder et Peter D. Vize, « Lunar Phase Modulates Circadian Gene Expression Cycles in the Broadcast Spawning Coral Acropora millepora », The Biological Bulletin, vol. 230, no 2, , p. 130–142 (ISSN 0006-3185, DOI 10.1086/BBLv230n2p130, lire en ligne, consulté le )
- O. Levy, L. Appelbaum, W. Leggat et Y. Gothlif, « Light-Responsive Cryptochromes from a Simple Multicellular Animal, the Coral Acropora millepora », Science, vol. 318, no 5849, , p. 467–470 (DOI 10.1126/science.1145432, lire en ligne, consulté le )
- Elias Levy, Golden Rabbitfish, (lire en ligne)
- (en) Akihiro Takemura, Md Saydur Rahman, Sigeo Nakamura et Young Ju Park, « Lunar cycles and reproductive activity in reef fishes with particular attention to rabbitfishes », Fish and Fisheries, vol. 5, no 4, , p. 317–328 (ISSN 1467-2979, DOI 10.1111/j.1467-2679.2004.00164.x, lire en ligne, consulté le )
- (en) Akihiro Takemura, Endang Sri Susilo, M. D. Saydur Rahman et Masaya Morita, « Perception and possible utilization of moonlight intensity for reproductive activities in a lunar-synchronized spawner, the golden rabbitfish », Journal of Experimental Zoology Part A: Comparative Experimental Biology, vol. 301A, no 10, , p. 844–851 (ISSN 1552-499X, DOI 10.1002/jez.a.105, lire en ligne, consulté le )
- (en-GB) H.-W. Korf, C. Schomerus et J. H. Stehle, « The Pineal Organ, Its Hormone Melatonin, and the Photoneuroendocrine System », Advances in Anatomy, Embryology and Cell Biology, (ISSN 0301-5556 et 2192-7065, DOI 10.1007/978-3-642-58932-4, lire en ligne, consulté le )
- (en) Akihiro Takemura, Satomi Ueda, Nanae Hiyakawa et Yoshiaki Nikaido, « A direct influence of moonlight intensity on changes in melatonin production by cultured pineal glands of the golden rabbitfish, Siganus guttatus », Journal of Pineal Research, vol. 40, no 3, , p. 236–241 (ISSN 1600-079X, DOI 10.1111/j.1600-079X.2005.00306.x, lire en ligne, consulté le )
- (en) B. B. P. Gupta, C. Haldar-Misra, M. Ghosh et J. P. Thapliyal, « Effect of melatonin on gonads, body weight, and luteinizing hormone (LH) dependent coloration of the Indian finch, Lal munia (Estrilda amandava) », General and Comparative Endocrinology, vol. 65, no 3, , p. 451–456 (ISSN 0016-6480, DOI 10.1016/0016-6480(87)90131-6, lire en ligne, consulté le )
- Martin Gühmann, English: Female epitoke of Platynereis dumerilii, (lire en ligne)
- (en) Juliane Zantke, Tomoko Ishikawa-Fujiwara, Enrique Arboleda et Claudia Lohs, « Circadian and Circalunar Clock Interactions in a Marine Annelid », Cell Reports, vol. 5, no 1, , p. 99–113 (PMID 24075994, PMCID PMC3913041, DOI 10.1016/j.celrep.2013.08.031, lire en ligne, consulté le )
- (en) Heinz -Dieter Franke, « On a clocklike mechanism timing lunar-rhythmic reproduction inTyposyllis prolifera (Polychaeta) », Journal of Comparative Physiology A, vol. 156, no 4, , p. 553–561 (ISSN 1432-1351, DOI 10.1007/BF00613979, lire en ligne, consulté le )
- HEINZ-DIETER FRANKE, « The Role of Light and Endogenous Factors in the Timing of the Reproductive Cycle ofTyposyllis proliferaand some other Polychaetes », American Zoologist, vol. 26, no 2, , p. 433–445 (ISSN 0003-1569, DOI 10.1093/icb/26.2.433, lire en ligne, consulté le )
- Auteur inconnuUnknown author, English: Diagram of Palolo Worm, date inconnueunknown date (lire en ligne)
- (en) Robert E. Markert, Betsy J. Markert et Nancy J. Vertrees, « Lunar Periodicity in Spawning and Luminescence in Odontosyllis Enopla », Ecology, vol. 42, no 2, , p. 414–415 (ISSN 1939-9170, DOI 10.2307/1932096, lire en ligne, consulté le )
- (en) Mercer R. Brugler, M. Teresa Aguado, Michael Tessler et Mark E. Siddall, « The transcriptome of the Bermuda fireworm Odontosyllis enopla (Annelida: Syllidae): A unique luciferase gene family and putative epitoky-related genes », PLOS ONE, vol. 13, no 8, , e0200944 (ISSN 1932-6203, PMID 30089107, PMCID PMC6082529, DOI 10.1371/journal.pone.0200944, lire en ligne, consulté le )
- (en) Hubert Caspers, « Beobachtungen über Lebensraum und Schwärmperiodizität des Palolowurmes Eunice viridis (Polychaeta, Eunicidae) I. Die tages- und mondzeitliche Konstanz des Schwärmens; Bibliographie », Internationale Revue der gesamten Hydrobiologie und Hydrographie, vol. 46, no 2, , p. 175–183 (ISSN 1522-2632, DOI 10.1002/iroh.19610460203, lire en ligne, consulté le )
- (en) H. Caspers, « Spawning periodicity and habitat of the palolo worm Eunice viridis (Polychaeta: Eunicidae) in the Samoan Islands », Marine Biology, vol. 79, no 3, , p. 229–236 (ISSN 1432-1793, DOI 10.1007/BF00393254, lire en ligne, consulté le )
- A. H. Haliday, English: Natural History Review, (lire en ligne)
- (en) Tobias S. Kaiser et David G. Heckel, « Genetic Architecture of Local Adaptation in Lunar and Diurnal Emergence Times of the Marine Midge Clunio marinus (Chironomidae, Diptera) », PLOS ONE, vol. 7, no 2, , e32092 (ISSN 1932-6203, PMID 22384150, PMCID PMC3285202, DOI 10.1371/journal.pone.0032092, lire en ligne, consulté le )
- (en) Dietrich Neumann, « Entrainment of a semilunar rhythm by simulated tidal cycles of mechanical disturbance », Journal of Experimental Marine Biology and Ecology, vol. 35, no 1, , p. 73–85 (ISSN 0022-0981, DOI 10.1016/0022-0981(78)90091-6, lire en ligne, consulté le )
- Ernest Naylor, Chronobiology of Marine Organisms, Cambridge University Press, (ISBN 978-0-521-76053-9, DOI 10.1017/cbo9780511803567, lire en ligne)
- (en) Dietrich Neumann, « Notizen: Photoperiodische Steuerung der 15-tägigen lunaren Metamorphose-Periodik von Clunio-Populationen (Diptera: Chironomidae) », Zeitschrift für Naturforschung B, vol. 20, no 8, , p. 818–819 (ISSN 1865-7117 et 0932-0776, DOI 10.1515/znb-1965-0825, lire en ligne, consulté le )
{kind=link}
{kind=link}
{kind=link}