Récepteur de type NOD
Les récepteurs de type NOD (NLR, nucleotide oligomerization domain receptors) sont des protéines cytoplasmiques pouvant avoir une fonction de récepteur de reconnaissance de motifs moléculaires.
Introduction
Le système immunitaire se compose de deux types de réponses : innée et adaptative. Les récepteurs NOD (Nucleotide-binding Oligomerization Domain proteins) font partie de la réponse immunitaire innée. Ils appartiennent à la grande famille des NLR (NOD-like receptors), avec les NALP et les NAID. La majorité de ces récepteurs est représentée par les récepteurs NOD1, NOD2, IPAF ou NLRC4. Ils reconnaissent des composants bactériens anormalement présents dans le cytoplasme des cellules, notamment le peptidoglycane (PGN) des bactéries GRAM positifs ou la flagelline bactérienne. Comme tous les PRR (pattern recognition receptors), ils participent à la reconnaissance du pathogène afin de déclencher divers signaux qui vont permettre d’engendrer une réponse anti-inflammatoire, résultant de l'immunité innée.
Phylogénie
Les gènes codant ces récepteurs sont très anciens et ont été conservés au cours de l'évolution; ils sont localisés sur les chromosomes 11, 16 et 19. On les retrouve chez les plantes, les insectes, et les mammifères. L'expression de NOD1 est observée dans toutes les cellules, tandis que NOD2 n'est exprimé que dans les cellules de l'immunité, les macrophages ou les cellules de Paneth.
Biologie de l'activation NOD
Architecture des domaines de NOD1 et NOD2
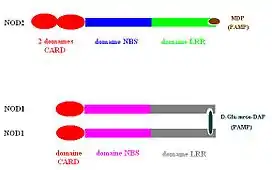
Les protéines NOD1 et NOD2 comprennent une série de leucine-rich repeats (LRRs) carboxy-terminaux, ainsi qu'un nucleotide-binding domain (NBD, aussi appelé domaine NOD) central, qui contient les sous-domaines NACHT, « winged helix », et « superhelical »[1].
Le domaine amino-terminal de NOD1 contient un caspase recruitment domain (CARD) unique, alors que chez NOD2 ce domaine contient des CARDs en tandem. Les domaines LRR des NODs sont impliqués dans la détection des ligands[2] - [3], et la reconnaissance du muramyldipeptide (MDP) par NOD2 ou de l'acide D‑glutamylmeso-diaminopimélique (iE-DAP) par NOD1 semble déclencher un changement conformationnel dans ces protéines, ce qui induit l'oligomérisation NBD-médiée des protéines NOD.
Caractéristiques structurales des motifs bactériens reconnus
NOD1 et NOD2 reconnaissent des PAMP (Pathogen-Associated Molecular Pattern) intracellulaires, plus précisément des constituants du peptidoglycane de la paroi bactérienne. NOD1 reconnaît le D-Glu-meso-DAP (acide gamma-D-glutamyl-meso-diaminopimelique), constituant des parois des bactéries à Gram négatif, tandis que NOD2 reconnaît le muramyl dipeptide (MDP, ou MurNAc-L-Ala-D-isoGln), présent dans les parois des bactéries à Gram négatif et à Gram positif.
Implication fonctionnelle des récepteurs NOD
L’activation des récepteurs NOD dans la réponse immunitaire innée permet la transmission d’un signal d’activation associé à la production de molécules effectrices.
Reconnaissance de PAMP
Lorsque le peptidoglycane ou un PAMP bactérien entre dans la cellule, celui-ci est reconnu par le domaine LRR des récepteurs NOD ce qui va entraîner un changement de conformation.
Transduction du signal
Le changement de conformation de NOD2 permet une oligomérisation de la molécule reconnue grâce au domaine NACHT. Sont ensuite recrutées la sérine-thréonine kinase RIP2 (RICK) et la caspase-9. Pour NOD1, deux récepteurs NOD1 sont nécessaires à la reconnaissance de PAMP. Il y a donc deux domaines CARD (un sur chaque récepteur) qui réagissent chacun avec un RIP2 : deux RIP2 interagissent avec deux CARD. Pour NOD2, ce récepteur possédant deux domaines CARD, va réagir avec deux RIP2. Dans les deux cas, le complexe CARD-RIP2 ainsi formé va permettre d'ubiquitiner les IKKγ (I-kappa B kinase-gamma) ou NEMO. Les IKK ubiquitinées vont entraîner la phosphorylation des sous-unités inhibitrices IκB qui sont liés au NF-κB (nuclear factor kappa-B) en les maintenant inactifs. Le changement de conformation des IκB libèrent alors les sous-unités actives du NF-κB qui deviennent compétentes. Les IκB sont dégradés par des protéasomes.
Expression de gènes impliqués dans l’inflammation
Les sous-unités p50 et p65 des NF-κB sont libérées et sont alors transloqués du cytoplasme vers le noyau. Ils vont pouvoir activer la transcription et la traduction de gènes codant des protéines de l’inflammation : cytokines pro- et anti-inflammatoires (IL-18, IL-1β pour NOD1…), peptides antibactériens.
Parallèlement, l’activation des récepteurs NOD1 activerait les mécanismes d’apoptose cellulaire, en coopération avec la caspase-9. Cet effet serait seulement potentialisé pour les récepteurs NOD2.
Régulation (non exhaustive)
- TRIP-6, LIM-domaine protéique interagit avec RIP2 afin de potentialiser l’activation de NOD1.
- CARD-6 interagit avec NOD1 et RIP2 en ralentissant l’activation des sous-unités p50 et p65 des NF-κB.
- TAK1 (transformating growth factor-bêta-activated kinase 1) interagit avec le domaine LRR de NOD2 pour inhiber l’activation des sous-unités p50 et p65 des NF-κB.
Pathologies associées
- Maladie de Crohn (une mutation sur le domaine LRR du gène de NOD2, ce qui modifie la reconnaissance du ligand)
- Asthme allergique (présence de NOD1 dans les poumons)
- Syndrome de Blau (mutation du gène NOD2 qui entraîne une hyper-activation de la voie NF-κB).
Notes et références
- (en) M. Proell, S.J. Riedl, J.H. Fritz, A.M. Rojas et R. Schwarzenbacher, « The Nod-Like Receptor (NLR) Family: A Tale of Similarities and Differences », PLoS ONE, vol. 3, no 4, , article no e2119 (DOI 10.1371/journal.pone.0002119)
- (en) S.E. Girardin, M.Jéhanno, D. Mengin-Lecreulx, P.J. Sansonetti, P.M. Alzari et D.J. Philpott, « Identification of the critical residues involved in peptidoglycan detection by Nod1. », The Journal of Biological Chemistry, American Society for Biochemistry and Molecular Biology, vol. 280, no 46, , p. 38648-38656 (DOI 10.1074/jbc.M509537200)
- (en) T. Tanabe, M. Chamaillard, Y. Ogura, L. Zhu, S. Qiu, J. Masumoto, P. Ghosh, A. Moran, M.M. Predergast, G. Tromp et al., « Regulatory regions and critical residues of NOD2 involved in muramyl dipeptide recognition », The EMBO Journal, vol. 23, no 7, , p. 1587-1597 (DOI 10.1038/sj.emboj.7600175)
Bibliographie
- Le Bourthis et al., « Roles of NODs in bacterial infection », Microbes and Infections, volume 9, 2007.
- Philpott et al., , The role of Toll-like receptors and Nod proteins in bacterial infection, Molecular Immunology volume 41, [PDF], consultable ici
- Cavaillon, 2010, The immune response to invasion: the ABCs – the innate immune system, extrait de l'article Early events in innate immunity in the recognition of microbial pathogens, Expert Opinion Biology Therapy
- Jeannin et al., juillet-, Biologie des récepteurs de l'immunité innée : applications cliniques et thérapeutiques, extrait du numéro 424 Revue Francophone des Laboratoires, [PDF]
- Jéru et al., 2011, Inflammasome et Interleukine 1, extrait de La Revue de médecine interne, volume 32, [PDF], consultable ici
- Tesnière et al., 2008, Immunité innée et sepsis : quand le danger vient de l'intérieur, [PDF] consultable ici
- Goutagn et al., , Dossier : les récepteurs de l'Immunité innée, extrait du numéro 368 Pour La Science
- Athman et al., 2004, Innate immunity via Toll-like receptors and Nod proteins, Current Opinion in Microbiology volume 7,[PDF], consultable ici
- Carneiro et al., , Innate immune recognition of microbes through Nod1 and Nod2:implications for disease, Microbe and infection volume 6,[PDF], consultable ici
Voir aussi
- NLRX1, une protéine de type NOD