Koinophilie
La koinophilie désigne une préférence pour les partenaires sexuels potentiels les plus communs comportementalement, ou ayant les caractères phénotypiques les plus habituels dans le groupe[1].
Un individu koinophilique évite de se reproduire avec un individu qui a des traits phénotypiques inhabituels, privilégiant la reproduction avec un partenaire ayant le phénotype le plus commun possible dans le milieu local.
Ainsi, un individu avec un trait phénotypique inhabituel sera moins attractif, pour un individu koinophilique, qu’un individu portant exclusivement des traits phénotypiques communs localement. Plus un individu a un nombre important de traits phénotypiques inhabituels, moins il est attractif pour tout individu koinophilique. En effet, même les individus portant des traits phénotypiques atypiques cherchent à se reproduire avec des individus au phénotype le plus commun possible, au niveau local[2].
Contexte génétique
Le génome est soumis à des mutations aléatoires, liées aux variations de l’environnement. Ces mutations vont conférer diverses caractéristiques physiologiques et morphologiques aux organismes qui les portent. Seuls les organismes possédant des mutations, leur conférant un avantage vis-à-vis de la population actuelle, vont être sélectionnés par sélection naturelle. Différents phénotypes sont donc présents dans la population, mais seuls les phénotypes les plus adaptatifs sont en grande proportion dans la population.
Lorsque l’on considère les organismes sexués, seules 50 % des caractéristiques génomiques et donc phénotypiques, de chaque parent sont transmises aux descendants. Dans un groupe sexué donné, le phénotype le plus transmis est celui qui est le plus fréquent, donc le plus commun et le plus adapté pour le milieu. Ainsi, la sélection naturelle va orienter les individus dans le sens d'une affinité pour ce type de partenaires, dit « communs ». C’est ce que Johan H. Koeslag a nommé la koinophilie[1].
Étymologie
Le terme koinophilie tire son origine du grec koinos qui signifie commun ou habituel et philos qui signifie aimer ou préférer.
Ce terme a été utilisé pour la première fois par le biologiste Johan Hans Koeslag dans les années 1990, à l'issue de ses recherches sur les stratégies évolutives. Ce terme a permis de mettre un nom sur le processus qu'il observait[3].
Causes évolutives
Évolution d’une population panmictique vers une population koinophilique
Dans les années 1990, J. H. Koeslag a réalisé une étude dont le but était de voir dans quelles circonstances un trait koinophilique initialement rare pouvait envahir une population qui se reproduisait jusque-là aléatoirement (panmixie).
Il a émis deux hypothèses, son hypothèse nulle étant que seul le hasard pourrait être responsable de l’invasion d’une population panmictique par la koinophilie. Son hypothèse alternative était que l’invasion de cette population par la koinophilie ne serait pas due au hasard.
Pour tester ses hypothèses, J. H. Koeslag a élaboré un modèle stochastique, où il pouvait faire varier différents paramètres, tels que le taux de mutation et la sélectivité maximale apportée par une mutation. Il a simulé une population de deux cents individus haploïdes, avec 50 % de mâles et 50 % de femelles. Chaque individu de cette population possédait cinquante traits phénotypiques, déterminés par un seul gène et, susceptibles de muter, au début de chaque génération. L’un de ces traits était celui de la préférence du partenaire sexuel, avec un allèle panmictique répandu à 95 % dans la population et un allèle koinophilique, présent à 5 % dans la population. À chaque épisode de reproduction, un couple d’individus ne pouvait donner qu’un seul descendant, panmictique ou koinophilique, selon les lois génétiques de Mendel. J.H. Koeslag a réalisé plusieurs séries. Au bout de 20 générations, il évaluait le niveau d’invasion de la koinophilie dans la population panmictique initiale[2].
Il a découvert que le facteur déterminant pour une telle invasion était la force de préférence du partenaire sexuel. En effet, la koinophilie tend à remplacer la panmixie toutes les fois où la préférence des traits phénotypiques communs est fortement exprimée, c’est-à-dire quand peu de mutations sont bénéfiques (1 % au lieu de 10 %) ou quand la fitness associée à ces mutations est faible (1.2 au lieu de 2.0). De plus, le choix koinophilique du partenaire remplace le choix aléatoire (panmixie) dès que les organismes sexuels possèdent une capacité suffisamment développée pour estimer si le partenaire sexuel potentiel a des traits phénotypiques habituels ou non[2].
Les populations koinophiliques, sujettes aux mutations à fitness aléatoire (dont, occasionnellement, des mutations bénéfiques), ont été démontrées comme ayant toujours un avantage considérable, immédiat mais temporaire, sur les populations panmictiques[2]. En effet, lorsque la koinophilie est rare, ceux qui la pratiquent cherchent à se reproduire avec des individus ayant des traits phénotypiques bénéfiques, adaptés à l'environnement local[4]. Ces derniers sont repérables de par leur augmentation progressive en fréquence dans la population. Cependant, quand la koinophilie devient plus commune, les individus qui la pratiquent ne se reproduisent plus qu’avec des individus avec un phénotype habituel et donc les nouvelles mutations bénéfiques ne s’établissent plus dans la population.
La stratégie de choix du partenaire koinophilique est donc une stratégie évolutivement stable par rapport à la stratégie panmixique. Néanmoins, comme la koinophilie entraîne une stagnation du phénotype (éliminant la variation), les populations panmixiques finissent par dépasser au bout d’un moment les populations koinophiliques en termes de fitness absolue[5].
Une population panmixique peut donc évoluer en population koinophilique, comme l’illustre le schéma, ci-dessous.

La koinophilie est donc un mécanisme qui permet d’expliquer en partie la sélection sexuelle. Ce mécanisme pourrait également expliquer pourquoi certaines espèces ne subissent aucun changement phénotypique durant de nombreuses années[2].
Reproduction sexuée et Fitness
Les organismes sexués, grâce à la reproduction et le brassage génétique associé, évitent la propagation de mutations délétères chez leurs descendants. Ainsi la reproduction sexuée prend en considération la fitness du partenaire car celui-ci peut transmettre 50 % de son génome à chaque descendant[1].
Les individus sont donc amenés à limiter leur choix de partenaire puisque leur but est de maximiser leur capacité à produire une descendance capable de se reproduire[6]. Tout l’enjeu est donc de choisir le partenaire possédant les « bons gènes » et d’éviter toute mutation susceptible d’être désavantageuse[5] - [7] - [6]. Pourtant il est presque impossible de prédire la fitness découlant d'une nouvelle mutation, et de savoir alors si elle sera bénéfique ou non pour l'individu. De façon générale, les mutations sont considérées comme étant délétères bien qu'il existe des mutations favorables[5]. Les individus sexués subissent donc une pression sélective dans le choix de leur partenaire. Le meilleur et seul indicateur est le fait que le partenaire présentant le phénotype le plus commun possible est le plus adapté dans son milieu et dans son groupe[1]. Ce choix sélectif va donc considérablement réduire les mutations du potentiel reproducteur et leur capacité à envahir la population[5].

Ce choix de partenaire peut discriminer les phénotypes mutants et de la même façon, peut discriminer plus généralement tout nouveau phénotype dans le groupe. Les individus vont chercher à éviter de s'accoupler avec tout partenaire ayant un phénotype inhabituel bien qu'il soit potentiellement bénéfique, neutre ou délétère[1]. On parle ici de la notion de préférence ; préférence pour les traits communs ou prépondérants dans le groupe[5] - [7] - [6].
Ainsi la part de mutations chez les descendants à travers un accouplement koinophilique est minimisée et le polymorphisme phénotypique est réduit[1]. De cette façon, lorsque la koinophilie évolue pour la première fois dans une population panmictique, chaque groupe koinophilique est composé d'individus ayant des phénotypes qui sont les plus adaptatifs.
.jpg.webp)
Une fois mis en place, un tel groupe koinophilique résisterait aux changements phénotypiques par mutation ou par immigration[1].
Chez les oiseaux, le choix du partenaire est souvent basé sur les dialectes du chant. Le dialecte correspond aux chants qui peuvent varier selon la région. Chez la mésange charbonnière (Parus major), les femelles évitent les mâles qui utilisent des chants rares, différents et non conformes au dialecte local. Ces mâles proviennent généralement d'endroits extérieurs au bois où les femelles sont nées ou des mutants de ce bois[1].
L’implication du système cognitif
La koinophilie agit sur le phénotype entier, et concerne notamment, le comportement social , la morphologie, la coloration, ou encore la fonction motrice. En effet, ce sont les caractéristiques qui ne peuvent manquer d’être remarquées par les partenaires potentiels. Bien que les individus d’une espèce se ressemblent, il est possible pour les congénères, de remarquer la moindre déviance à la norme. Cette reconnaissance entre individus fait intervenir le système cognitif[6] - [1].

Les premières manifestations de ce phénotype le plus commun auraient favorisé l'évolution des mécanismes internes à chaque individu, de distinctions de ce phénotype commun parmi tous les autres dans une communauté (analyse des fréquences alléliques) . La sélection naturelle associée à la réplication de l'ADN rend toujours rares les traits (caractères phénotypiques) inadaptés, (car ils apportent un avantage moindre en termes de fitness) contrairement aux traits communs. C’est pourquoi l'analyse des fréquences alléliques constitue un bon indicateur du bon type de partenaire et des partenaires à éviter[1]. Selon la théorie de l’attribut du groupe, les traits les moins adaptatifs sont concentrés à un faible niveau sur le spectre des fréquences alléliques, et les traits adaptatifs, à un niveau supérieur[5].
Ce système permet alors d'améliorer et d’affiner le processus de sélection du partenaire et de fiabiliser la préférence dans le choix du partenaire, car, ce qui est commun, répandu ne peut être trompeur et faux[6].
Conséquences évolutives de la koinophilie
Isolement reproductif
J. H. Koeslag a modélisé l'évolution d'une population koinophilique selon la distribution de deux traits, et selon la répartition géographique des individus[7]. Il a observé qu'un isolement reproductif correspond à un mécanisme empêchant ou limitant fortement l’hybridation de deux populations vivant dans la même région, même lorsque celles-ci sont étroitement apparentées. Ainsi, l'isolement reproductif pré-zygotique fait obstacle aux croisements avant fertilisation, de par la présence d'une barrière géographique (montagne, falaise...) entre deux populations ou des périodes de reproduction qui diffèrent entre populations. L'isolement reproductif post-zygotique empêche quant à lui les descendants hybrides de survivre ou de se reproduire.

Cet isolement reproductif va conduire à la formation de regroupements entre individus koinophiliques présentant un phénotype similaire ou proche[7]. La fitness des groupes les plus représentés en nombre va être supérieure à celle des petits groupes. Breinstein et ses collègues vont définir cela comme étant le coût de la rareté. En effet, les grands groupes vont encore s'agrandir aux dépens des petits groupes qui vont continuer à diminuer[7] - [1]. En fait, les individus d'un groupe koinophilique plus grand trouveront un partenaire plus facilement que les individus d'un plus petit groupe[1] - [7]. Par conséquent, au bout de plusieurs générations, seuls quelques groupes koinophiliques, distincts du point de vue de leur phénotype, vont persister et demeurer inchangés face aux changements environnementaux[1]. Les autres groupes, possédant un phénotype intermédiaire vont s'éteindre, formant des lacunes phénotypiques entre les groupes restants[5] - [7].
Dans ce contexte, la koinophilie apparaît comme une première étape au développement des barrières à l'hybridation. Ce type de choix du partenaire peut être considéré comme un moteur de la spéciation.

Les papillons Euphydryas editha de Jasper Ridge aux États-Unis, se répartissent en trois populations sexuellement séparées, bien qu'elles ne soient confrontées à aucun obstacle pour un accouplement aléatoire[1].
L'isolement sexuel de ces trois populations semble être maintenu par koinophilie. Le comportement d'évitement des phénotypes non communs à chaque population a été mutuel[1].
La koinophilie semble avoir gardé les papillons de Jasper Ridge séparés par des différences extrêmement subtiles, principalement induites par l'environnement des populations. Ainsi, les papillons choisissent leurs partenaires selon les habitudes alimentaires ou l'odeur des individus, qui doivent être les plus proches de celles du type habituel retrouvé dans la zone géographique où ils ont été élevés[1].
La stabilité des espèces
La Koinophilie pourrait aussi expliquer la plupart des changements phénotypiques évolutifs qui sont conservés chez les animaux et le fait que ces évolutions aient été réalisées par des modifications mineures et progressives[5]. En effet, une fois fixée, la koinophilie limite la capacité d'évolution des phénotypes et maintient le phénotype dominant dans une espèce donnée. Ainsi, la fréquence allélique dominante dans la population reste stable, et résiste aux changements évolutifs, malgré les mutations cryptiques (ie qui n’affectent pas le comportement) accumulées[5]. C’est pourquoi on peut observer de longues périodes de stagnation évolutive entre les évènements de spéciation ou apparition d'une nouvelle espèce du fait d'un haut degré intraspécifique d'uniformité phénotypique[6].
Le maintien du sexe
J.H. Koeslag[8] a tenté d’expliquer comment la reproduction sexuée s’est maintenue au cours de l’évolution. Pour cela il a utilisé un modèle avec une population initiale composée de différentes proportions d'individus sexués et asexués. Il a ensuite utilisé un modèle mathématique informatique afin de déterminer, au bout de plusieurs générations, quelles étaient les proportions d'asexués et de sexués qui se maintenaient dans la population finale[8]. Durant une génération plusieurs phases s'enchaînent avant d'aboutir à la nouvelle génération[9] :
- Mutation ⇒ Sélection ⇒ Rencontre entre deux partenaires (uniquement pour les individus sexué) ⇒ reproduction[9].
Dans un premier temps, il s'est focalisé sur une population constituée d'individus sexués et asexués. Après plusieurs générations, les individus asexués envahissent rapidement la population au détriment des individus sexués qui disparaissent. Le résultat est le même lorsque l'on fait varier le taux de mutation, lorsque l'on insert des mutations avantageuses ou lorsque l'on change la fitness moyenne des mutants. Ces résultats montrent que les individus sexués n’arrivent pas à se maintenir au sein de la population finale, mais n’expliquent pas pourquoi les individus sexués restent présents dans la nature[8] - [9].
Or, d'après J. H. Koeslag, l'individu sexué doit se reproduire uniquement avec un partenaire sexuel qui maximise sa fitness. À cause de cette pression de sélection, l'individu sexué koinophilique doit être capable de distinguer les traits avantageux de ses partenaires sexuels[1]. Généralement dans une population les traits inadaptés ont tendance à être représentés en faible proportion alors que les traits adaptés ont tendance à être représentés en forte proportion[1] ; de cette manière, les individus sont sûrs de pouvoir éviter de se reproduire avec des individus ayant des traits phénotypiques rares ou inhabituels, c’est dans ce cas qu'intervient la koinophilie[8] - [9].
Ainsi, Koeslag a essayé de vérifier si les populations sexuées arrivaient à se maintenir grâce à la koinophilie. Il intègre la koinophilie dans son modèle comme étant une probabilité de reproduction entre un individu koinophilique et un autre individu dépendant du niveau d'attractivité et de la fitness de cet individu, l'attractivité et la fitness étant proportionnelles[8] - [9]. Il expérimente une population initiale constituée d'individus sexués koinophiliques et asexués. Selon son modèle informatique et après plusieurs générations les individus asexués n'envahissent pas la population sous certaines conditions.
Dans ces modèles, Koeslag a constaté qu’un fort taux de mutation entraînait une diminution de la proportion des individus asexués, pour obtenir finalement une population constituée uniquement d’individus sexués koinophiliques. Plus le taux de mutation augmente et plus la population d’asexués n’arrive pas à se maintenir. Dans le cas contraire l’effet est inversé, une diminution du taux de mutation entraîne la fixation des individus asexués au détriment des individus sexués koinophiliques[8].
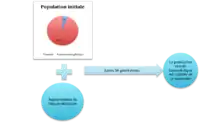
Selon que la mutation soit bénéfique ou pas, les résultats sont différents. Quand les mutations ne sont pas bénéfiques les asexués n’arrivent pas à envahir les sexués koinophiliques qui se maintiennent au sein de la population finale. Inversement, une augmentation des mutations bénéfiques mène au maintien des asexués aux dépens des sexués koinophiliques[8].
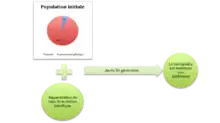
En considérant la valeur sélective (fitness) moyenne des mutants, une augmentation de la valeur sélective moyenne des mutants permet à la population sexuée koinophilique de se maintenir après plusieurs générations. Les mutants arrivent donc mieux à se reproduire et les asexués ont du mal à maintenir la population. Par contre, une diminution de la valeur sélective moyenne des mutants entraîne le maintien de la population asexuée, les individus koinophiliques ont du mal à subsister[8].
_moyenne_des_mutants.png.webp)
Rappelons que dans les mêmes conditions, les individus sexués (non koinophilique) n’arrivaient pas à se maintenir au sein de la population finale[8].
Par conséquent, les individus sexués koinophiliques ont une puissante capacité de rejet d'erreur génétique qui leur confère un avantage face aux individus asexués et leur permet de se maintenir dans la population. La koinophilie est donc capable de stabiliser les populations sexuées face à celles asexuées, malgré le coût de production élevé pour les individus sexués[9].
L’avantage de la koinophilie réside clairement dans sa capacité à résister à l’accumulation de mutations légèrement désavantageuses dans sa descendance. Elle permet la formation de groupe avec un morphisme qui lui est propre et qui résiste à l’hybridation avec d’autres groupes en créant des lacunes morphologiques entre les populations[8] - [9].

Cependant, lorsque le choix du partenaire est limité par la faible densité de population par exemple dans les environnements arides, l'efficacité de la koinophilie comme dispositif de rejet d’erreur génétique peut être gravement compromise.
De plus, l'invasion d'hétérozygotes présentant des allèles dominant dans une population koinophilique entraîne un déclin progressif de l'attraction et de la présence du trait commun après plusieurs générations. L’idée qu'« un bon partenaire est celui qui présente les meilleurs traits (souvent rares) » intervient au détriment de la koinophilie qui dit qu'« un bon partenaire possède les traits communs ». Deux issues sont alors possible, soit les individus se reproduisent malgré l'existence d'un polymorphisme au sein de la population (la koinophilie a disparu), soit d’éventuels indicateurs indirects de fitness font leur apparition, comme un ornement sexuel maladroit[10].
Les limites de ces modèles

Catégoriser les individus comme étant sexués ou asexués ne permet pas d’en tirer les bonnes conclusions. En effet, de nombreuses espèces peuvent être définies par le terme "sexué", il en est de même pour les individus asexués. Il peut exister différente forme de sélection sexuelle selon l’espèce sexuée considérée[10]. De plus le taux de mutations peut être différent selon que l’individu soit sexué ou asexué, et particulièrement selon l’espèce. Généralement, le taux de mutation par paire de base est plus élevé chez les asexués que chez les sexués sauf dans des cas particuliers[11] - [12].
Certaines espèces peuvent faire de la reproduction sexuée et asexué selon leur cycle de vie ou le temps, c’est le cas du Dragon de Komodo. C’est dans un zoo qu’une femelle Varanus komodoeusis ou Dragon de komodo va donner naissance à cinq jeunes dragons par parthénogenèse, sans qu’elle n’ait eu de contact avec un mâle, ce qui est un événement assez rare[13].
Ces modèles ne trouvent pas d’exemple concret dans la nature.
Implication de la koinophilie dans la coopération
_lat._B._Walker.jpg.webp)
L’une des conséquences de la koinophilie est de promouvoir l’accumulation d’avantages sélectifs au niveau du groupe, qui ne sont pas forcément bénéfiques pour l’individu (Koeslag, 1994). Le dilemme du prisonnier, énoncé par Albert W. Tucker en 1950, caractérise une situation où deux joueurs doivent choisir de coopérer ou de trahir l'autre sur un jeu qui ne dure qu'un tour. Ainsi, si les deux joueurs choisissent de coopérer, leur bénéfice sera maximal. Si seul un des deux joueurs coopère, il sera fortement pénalisé tandis que l'autre joueur aura un bénéfice encore plus grand. Cependant, si les deux joueurs se trahissent, le résultat leur sera moins favorable que si les deux joueurs avaient coopéré. Cette théorie illustre donc l'avantage pour deux individus de coopérer afin de maximiser leurs bénéfices. La koinophilie pousse à la coopération au sein du groupe et permet donc de maximiser la fitness du groupe entier[14].
Les études de Lande (1981) et Kirkpatrick (1982) ont montré que la koinophilie ne s’appliquait pas seulement à des critères phénotypiques mais qu’elle concernait aussi les comportements et surtout les interactions sociales des individus au sein de la population[12] - [10]. Ils ont montré que la koinophilie stabilise les dynamiques de la population en favorisant tout ce qui conduit à l’élaboration d’une stratégie commune. Si la koinophilie est négligée lors d’une étude de dynamique populationnelle, aucune stratégie évolutivement stable n’est trouvée. Dans les populations koinophiliques, la stratégie individuelle laisse place à une stratégie de groupe. Le groupe est dit d'« accumulateurs d’adaptations » car la coopération entre les individus permet de maximiser la fitness du groupe. En effet, les groupes coopératifs sont plus efficaces que les individus égoïstes car ils exploitent de façon plus efficace leur environnement[15]. Cependant, cette maximisation de la fitness ne fonctionne qu’à l’échelle du groupe. Ainsi, si un individu quitte son groupe d’origine il perdra tous les avantages de sa stratégie de coopération puisque cette dernière ne s’applique pas à l’individu mais au groupe. En effet, cette stratégie évolutivement stable est héritée de génération en génération par le groupe et non par l’individu.
Les groupes coopératifs sont donc plus avantagés par rapport aux individus égoïstes. Pourtant, les individus égoïstes possèdent un niveau de fitness individuelle plus élevé que les individus de ces groupes[15] - [3]. En effet, si on considère un individu égoïste, dans un groupe présentant une stratégie coopérative pour la chasse, ne participant donc pas à la chasse comme les autres individus du groupe, et profitant tout de même de la nourriture rapportée, il paraît évident que celui-ci sera plus avantagé qu’un individu qui coopère. Cet individu égoïste aura probablement un taux de survie et un taux de reproduction plus important que les autres membres du groupe. Cependant, ces individus égoïstes ne peuvent pas entrer en compétition avec des groupes présentant une bonne coopération[15] - [3]. En effet, si cet individu égoïste est considéré comme inadapté pour le groupe par ses congénères, parce qu’il ne possède pas les caractéristiques comportementales standards du groupe, il sera rejeté. Il devient alors évident qu'il serait plus avantageux pour chaque individu du groupe de coopérer.
La koinophilie stabilise les phénotypes comportementaux et les préférences des femelles, ce qui est comparable au modèle de Lande et Kirkpatrick sur la stabilisation des ornements chez les mâles. La coopération dans un groupe se ferait en plusieurs étapes, en commençant pas un caractère bien spécifique, exemple : la recherche de nourriture. Cette caractéristique avantageuse évolue et engendre une coopération entre les individus puis s’étend ensuite à d’autres caractères tels que le cri d’alarme ou autre, c’est ainsi qu’un groupe devient coopératif, par étapes. Cela soulève les capacités d’imitation et d’apprentissage chez les animaux avec une forte plasticité comportementale. Il est difficile de séparer la koinophilie de la coopération. Puisque sans coopération, la koinophilie serait vite éliminée par transfuge et que sans koinophilie, la coopération disparaîtrait au bout de seulement six générations. La koinophilie est définie comme une « force évolutive agressive » de la coopération[12].
La koinophilie demande de grandes capacités mentales de traitement de l’information ainsi que de mémoire car il est nécessaire d’analyser à la fois les comportements du groupe dans différentes circonstances mais aussi le comportement du partenaire potentiel. Il y a toute une étude sur les mécanismes physiologiques mémoire-sensoriels en cours de réalisation.
Du point de vue évolutif, la koinophilie permet donc au comportement coopératif de stabiliser les groupes[15].
Chez certains oiseaux nicheurs tels que le Geai à gorge blanche (Aphelocoma coerulescens) les individus qui ne sont pas des ‘aides’ pour le groupe (construction des nids, protection du territoire, nourrissage des petits) s'accouplent rarement . En effet, les oiseaux qui ont une certaine capacité à se reconnaître, choisissent de ne pas s'accoupler avec des partenaires ayant ce comportement non conforme[15].
Valeur de la théorie de J.H Koeslag
J.H. Koeslag est le principal investigateur des études au sujet de la koinophilie : il a publié de nombreux articles s’appuyant sur des modèles stochastiques pour illustrer ce phénomène. Cependant, la communauté scientifique n’ayant pas trouvé d’exemple que cette théorie s’appliquait au vivant, elle l’a peu à peu laissé tomber dans l’oubli[16]. Il semblerait en effet, que la plupart des espèces orienterait leur choix du partenaire sexuel en fonction du critère d’adaptabilité. En fait, plus un partenaire sexuel potentiel va être adapté à son environnement, plus il va trouver facilement un partenaire. Dans un contexte comme celui-ci, la koinophilie ne pourrait jamais envahir cette population. En effet, la koinophilie ne garantit pas à 100 % le choix d’un bon partenaire, tandis que le choix du partenaire le plus adapté assure cela.
De plus, Koeslag attribue à la koinophilie de nombreux bénéfices qui ne lui sont pas dus. Par exemple, il certifie que les mutations délétères sont éradiquées plus rapidement dans une population koinophilique que dans toute autre population. Néanmoins, quelle que soit la stratégie de choix du partenaire sexuel, une mutation délétère est toujours très rapidement éliminée de la population du fait que l’individu la portant se reproduit moins que les autres car il est mal adapté.
D’autre part, contrairement à ce que prétend J. H. Koeslag, la koinophilie n’apporte pas de maintien de la sexualité au sein d’une population. En effet, s’il existe un nombre suffisant de mutations bénéfiques dans une population, alors quel que soit le type de choix du partenaire sexuel, la population asexuée se maintiendra au détriment de la population de sexuée.
Ainsi la koinophilie reste une théorie assez controversée et limitée en termes d’application[16].
Notes et références
- Johan H. Koeslag, Koinophilia groups sexual creatures into species, promotes stasis, and stabilizes social behaviour, Journal of theoretical biology, 1990, vol. 144, no 1, p. 15-35.
- J. H. Koeslag, Koinophilia replaces random mating in populations subject to mutations with randomly varying fitnesses, Journal of theoretical biology, 1994, vol. 171, no 3, p. 341-345.
- « Johan (Hans) Koeslag », sur Old Cambrian Society, (consulté le )
- (en) Johan H. Koeslag, Koinophilia groups sexual creatures into species, promotes stasis, and stabilizes social behaviour, Journal of theoretical biology, 1990, vol. 144, no 1, p. 15-35.
- (en) Johan H. Koeslag et Peter D. Koeslag, Koinophilia, Journal of theoretical biology, 1994, vol. 167, no 1, p. 55-65.
- M. K. Unnikrishnan, Koinophilia revisited: the evolutionary link between mate selection and face recognition, Current Science(Bangalore), 2012, vol. 102, no 4, p. 563-570.
- Johan H. Koeslag, On the engine of speciation, Journal of theoretical biology, 1995, vol. 177, no 4, p. 401-409.
- (en) J. H. Koeslag et P. D. Koeslag, Evolutionarily stable meiotic sex, Journal of Heredity, 1993, vol. 84, no 5, p. 396-399.
- (en) Peter D. Koeslag et Johan H. Koeslag, Koinophilia stabilizes bi-gender sexual reproduction against asex in an unchanging environment, Journal of theoretical biology, 1994, vol. 166, no 3, p. 251-260.
- (en) Malte Andersson et Leigh W. Simmons, Sexual selection and mate choice, Trends in Ecology & Evolution, 2006, vol. 21, no 6, p. 296-302.
- (en) Paul D. Sniegowski, Philip J. Gerrish, Toby Johnson, et al., The evolution of mutation rates: separating causes from consequences, Bioessays, 2000, vol. 22, no 12, p. 1057-1066.
- (en) John W. Drake, Brian Charlesworth, Deborah Charlesworth, et al., Rates of spontaneous mutation, Genetics, 1998, vol. 148, no 4, p. 1667-1686.
- (en) Phillip C. Watts, Kevin R. Buley, Stephanie Sanderson, et al., Parthenogenesis in Komodo dragons, Nature, 2006, vol. 444, no 7122, p. 1021-1022.
- (en) Johan H. Koeslag et Elmarie Treblanche, Evolution of cooperation: cooperation defeats defection in the cornfield model, Journal of theoretical biology, 2003, vol. 224, no 3, p. 399-410.
- (en) Johan H. Koeslag, Sex, the prisoner's dilemma game, and the evolutionary inevitability of cooperation, Journal of theoretical biology, 1997, vol. 189, no 1, p. 53-61.
- (en)MATHEW, Geetha. Koinophilia and the exception reporting model of face recognition: Ill-fated solitary neologisms?. Current Science, 2012, vol. 102, no 10, p. 1354.