Glande clypéale
La glande clypéale, appelée aussi glande acronale, est un organe sécrétant, propre aux mâles du genre des Argyrodes, araignées appartenant à la famille des Theridiidae[1].
Elle se situe dans la partie antérieure du céphalothorax - appelé aussi prosoma - et joue un rôle capital lors de l'accouplement[2].
Introduction
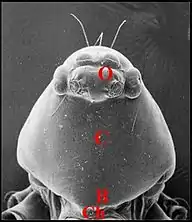
Le clypéus des Araignées est un territoire plus ou moins étendu du céphalothorax correspondant à la partie toute antérieure de sa face dorsale (carapace ou bouclier) (Fig.1). Encore appelé bandeau, il sépare les yeux (O) du bord frontal (B) surplombant les chélicères (Ch). Chez les mâles de certaines araignées, essentiellement les Theridiidae du genre Argyrodes, et, en extrapolant à certaines Linyphiidae (Mynoglenes)(Fig.2) ou Erigoninae, il est en relation
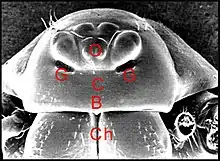
profonde avec un épiderme dont les cellules se sont différenciées en organes tégumentaires sécréteurs plus ou moins complexes, les glandes dites clypéales, sensu stricto chez les premiers, et au sens large, par abus de langage, dans les deux autres groupes.
Localisation
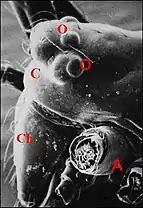
Cet organe prosomatique insolite, découvert par André Lopez chez Argyrodes zonatus de Madagascar est propre aux mâles du même genre , sans équivalent chez leurs femelles et remarquable par sa localisation[1] - [3]. Il siège en effet dans la région céphalique du corps de l'Argyrodes mâle, plus précisément dans la partie toute antérieure du prosoma où ses rapports avec le clypeus avaient d'abord inspiré son nom de glande clypéale[1], consacré par l’usage et étendu à d’autres familles d’Araignées. En fait, cette appellation globale ne paraît plus justifiée car chez certaines espèces du type acuminé, la glande n’a plus, au moins en partie, de relation directe avec le bandeau. Sa présence est étroitement liée à celle d’un dimorphisme sexuel céphalothoracique très marqué.
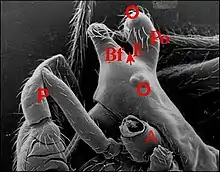
Rapports avec le dimorphisme sexuel
Alors que le prosoma des femelles a un aspect banal et assez uniforme (Fig. 3), celui des mâles (Fig.4) présente une morphologie déjà utilisée dans la classification puisque certains auteurs (Pickard-Cambridge, Exline & Levi, Chrysanthus) ont eu recours aux détails du céphalothorax dans un but systématique.
Le dimorphisme sexuel des Argyrodes

Son aspect extraordinaire, bien visible au microscope électronique (M.E.B) dans la région du clypeus , associe des saillies, bosses ou protubérances diverses et des dépressions, échancrures ou sillons plus ou moins garnis de poils. Leur ensemble confine parfois à l'extravagance (Fig.4)(voir d'autres aspects ou morphotypes dans l'article Argyrodes : fig.5 à 9). L’étude au M.E.B[4] montre aussi l’aspect du tégument (lisse ou garni de reliefs en écailles), l’existence de saillies accessoires plus ou moins tronconiques (en chicots chez Argyrodes amplifrons) tendant à fermer l'échancrure ou le sillon, la structure des poils et surtout, l’existence d’orifices. Ces derniers sont béants, sans bourrelet complet, parfois circonscrits par deux lèvres, non surélevés, le plus souvent de niveau avec la surface tégumentaire,
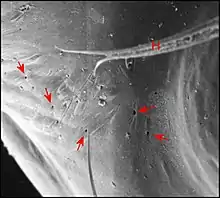
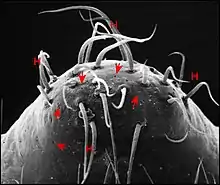
rarement dans le fond de fentes étroites en boutonnières (Argyrodes fissifrontella, Seychelles)[5]. Ils peuvent être dispersés sans ordre apparent (type acuminé) ou réunis en deux groupes de 2 à 5 orifices chacun, symétriques par rapport au plan sagittal (type rostré). Dans tous les cas, ils sont nettement séparés des poils, y compris chez l’espèce néozélandaise Argyrodes antipodiana où ils ne s’ouvrent pas dans leur alvéole basale, bien que Whitehouse[6], malgré son utilisation du M.E.B., ait affirmé le contraire par manque d’étude histologique. Ces orifices sont précisément les pores excréteurs de la glande clypéale sous-jacente déjà perceptible par transparence dans des céphalothorax éclaircis artificiellement (Fig.5).
Les divers morphotypes
Ces reliefs ont été initialement considérés comme "sans utilité" par Berland (1932), puis illustrés par Millot qui ne leur attribue aucune signification(1949), reconnus enfin par Legendre (1960)[2] comme jouant un rôle de premier plan dans l'accouplement. Suivant leur aspect général et leur disposition[4] - [5] - [7]) il a été proposé de rattacher les mâles d’ Argyrodes à six types morphologiques bien distinctsprésentés en détails dans l'article Argyrodes.avec les figures référencées ci-après: types "acuminé" (Argyrodes cognatus)(fig.5 in Argyrodes), "rostré" (Argyrodes zonatus, Argyrodes argyrodes, A. elevatus, A. nephilae et A. rostratus nephilae)(fig.2 in Argyrodes), "nasuté" (Argyrodes borbonicus, A. nasutus) (fig. 6 in Argyrodes), "prognathe" (Argyrodes benedicti Lopez), de Guyane, A.cochleaforma, A. sullana, A.atopus, A. proboscifer) (fig.7 in Argyrodes), "lippu" (Argyrodes amplifrons et A.ululans de Guyane française) (fig.8 in Argyrodes) et "camard" (Argyrodes caudatus, A.cancellatus, tous deux des Antilles françaises) (fig.9 in Argyrodes).
Description de la glande clypéale ou acronale
Elle est déjà perceptible par transparence dans des céphalothorax éclaircis artificiellement (Fig. 5 et Fig. 10 in Argyrodes). Sa connaissance est fondée sur les études histologiques et essentiellement sur les recherches au microscope électronique à transmission: Argyrodes argyrodes de type rostré[8] - [9] et Argyrodes cognatus, de type acuminé[7].
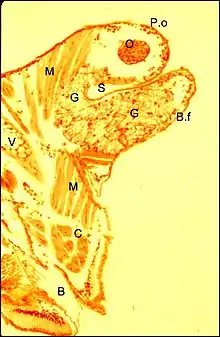
Elles en montrent respectivement la structure d’ensemble et la structure fine de l’unité glandulaire. De plus, les coupes histologiques horizontales révèlent que la glande clypéale n'est pas un organe impair mais bilatéral, car formé par deux parties distinctes, droite et gauche, plus ou moins asymétriques, et la microscopie électronique à balayage individualise une bipartition des orifices excréteurs (..). Toutefois, chez les Argyrodes cognatus et fissifrontella, il ne semble exister qu'un seul massif impair, à pores ouverts sur toute la surface de la "tourelle" et de l'échancrure (Fig.9,10) .
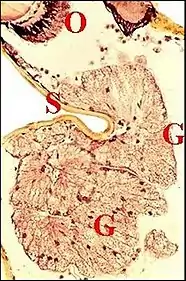
Structure histologique
Dans le type "rostré" que présente Argyrodes argyrodes (Fig.5), A. zonatus (Fig.4,6), A.elevatus, et A.nephilae (Fig.7), le type "lippu" d'Argyrodes ululans (Fig.8) et le type "camard" (Argyrodes caudatus, A.cancellatus)(Fig.9), un canal collecteur principal aboutit à chacun des pores.
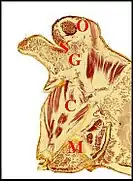
Il résulte de la confluence de canalicules excréteurs de deuxième ordre; ces derniers sont eux-mêmes formés par l’union de canalicules excréteurs de premier ordre, chacun drainant une cellule sécrétrice ou adénocyte et constituant avec elle l’unité glandulaire.
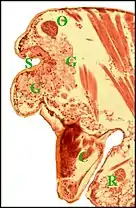
Les canaux excréteurs de tous ordres sont entourés par des cellules canalaires. L'ensemble de tous ces canaux constitue un appareil cuticulaire s’étendant des portions réceptrices aux pores de surface. Les adénocytes se groupent par 5 ou 6 en un petit lobule assez bien individualisé. L'ensemble des lobules constitue l’un des deux massifs compacts incurvés (Fig.6,7,8) ou des cordons sinueux (fig.9,10,11) dont est formée la glande[1] - [3] - [4]). Chaque massif ou cordon d'adénocytes aboutit donc séparément à l’un des deux groupes de pores du sillon ou échancrure par le système des canaux interposés (glandes arborescentes). Toutefois, chez l’espèce Argyrodes fissifrontella (Seychelles), de morphotype rostré, l’ensemble de l’organe ne paraît pas bilobé mais impair (Fig.10); ses unités glandulaires sont isolées ou en lobules lâches; leurs canaux ne se limitent pas à la bosse frontale mais s’ouvrent également sur la protubérance oculaire[5].
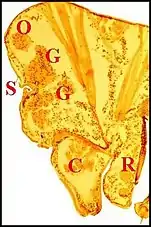
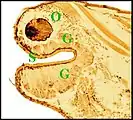
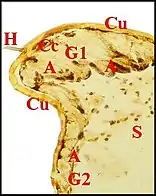
Dans le type "acuminé" (Argyrodes cognatus)(Fig.13) un massif glandulaire également impair
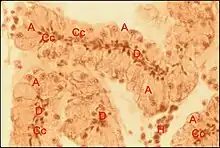
occupe la protubérance sus-oculaire en tourelle (turret gland) et se prolonge en languette sous le tégument clypéal[7]. Il est formé par des unités juxtaposées bien séparées ou à disposition lobulaire peu marquée, chacune d’elles comprenant un adénocyte, une seule cellule canalaire et un canal excréteur de 1er ordre, aboutissant directement à un pore, parfois après s’être fusionné avec un voisin juste avant cette terminaison. Comme chez A.fissifrontella, il n’existe pas de système excréteur ramifié (glandes "simples").
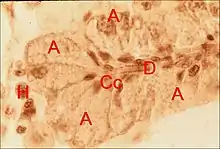
Ultrastructure
Son étude permet seule de réconstituer l'organe avec ses unités glandulaires.
Unité glandulaire
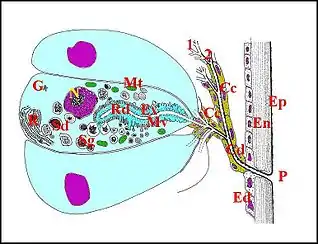
Elle représente une structure anatomo-fonctionnelle de base, autonome, bien individualisée, constituée par une cellule glandulaire ou adénocyte et son appareil cuticulaire comportant deux portions, réceptrice et conductrice (Schéma).
Adénocyte
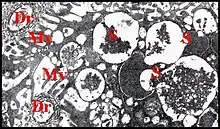
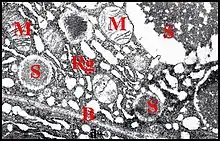
L’adénocyte est pyramidal ou piriforme, long en moyenne de 65 µm, pourvu d’un noyau vésiculeux généralement basal, n’excédant pas 9 µm, et d'un cytoplasme assez compact et basophile en périphérie, d’aspect vacuolisé et spongieux dans la majeure partie de son étendue, sans sécrétion figurée. Il renferme un canalicule axial surtout visible en coupe transversale, et qu’entoure une zone arrondie en réservoir acidophile, faiblement PAS +.
Il est enveloppé par une gaine de lame basale le séparant du sinus hémolymphatique adjacent. Sa membrane plasmique ne présente pas de dispositifs (replis) en augmentant la surface au niveau du pôle externe (basal), qui est régulièrement arrondi, et des faces latérales, qui sont à peu près planes. En revanche, elle pénètre très profondément dans le pôle apical qui présente ainsi une invagination de l’espace extracellulaire en cul de sac ou doigt de gant. Ce réservoir occupe en longueur la moitié de l’adénocyte, est sinueux, bordé par des microvillosités (considérées d’abord à tort comme des canaux intra-cytoplasmiques !)[8] et renferme un canalicule récepteur en position axiale (Schéma). Les microvillosités (Fig.) sont courtes et espacées près de l’apex, plus longues, plus nombreuses et resserrées dans sa partie distale, présentent des flexuosités et sont soutenues par du cytosquelette (quelques microtubules, surtout des filaments d’actine réunis en faisceaux qui s'ancrent sur une densification apicale).
Le noyau est plus ou moins déformé par les grains de sécrétion, renferme un nucléole volumineux et une chromatine finement grenue, dispersée, ou réunie en petites mottes éparses.Les organites subcellulaires sont des mitochondries ovoïdes dispersées dans le hyaloplasme, des ribosomes remarquablement nombreux et surtout, un réticulum endoplasmique extrêmement développé et de type rugueux. Il se présente sous la forme de tubules irréguliers, surtout localisés en périphérie, au voisinage du plasmalemme, et de citernes, très variables dans leur aspect et leurs dimensions, occupant surtout le reste de la cellule. Les empilements membranaires (“dictyosomes”) qui caractérisent en général un appareil de Golgi typique paraissent absents.
En revanche, une sécrétion abondante est élaborée dans certains adénocytes. Elle se présente sous la forme de grains (Fig.) contenus dans des vésicules semblant issues du réticulum qui les entoure : les unes renferment un matériel peu dense, floculant, irrégulier et grossissent par confluence ; les autres, non coalescentes, ont un contenu plus opaque, homogène et subsphérique (Fig.). Les mêmes vésicules peuvent fusionner et extrudent leur contenu entre les bases des microvillosités. Cette substance, finement grenue, pénètre ainsi dans l' espace extracellulaire, autour du canalicule récepteur.
Canalicule récepteur (portion réceptrice)
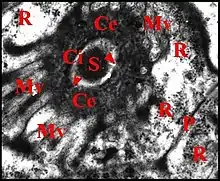
Il présente une lumière de 0,5 µ de diamètre et une paroi (épaisseur : 0,2 à 0,3 µ) comprenant deux couches : couche interne entourant la lumière, relativement homogène, perforée, et couche externe plus épaisse, lâche, d’aspect vermiculé, formée par des strates discontinues grossièrement concentriques se disposant en filtre (sieve) (Fig.). Le matériel sécrétoire finement grenu ayant gagné l' espace extracellulaire se retrouve dans les interstices de ce filtre, puis rejoint la lumière par les pores de la couche interne et s’y accumule.
Le canalicule récepteur entouré par les microvillosités correspond à un appareil terminal (end apparatus) typique (Schéma, Fig.). Il s’abouche à la portion conductrice qui est dans son prolongement direct.
Portion conductrice
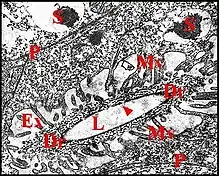
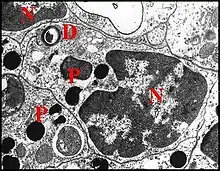
Difficilement perceptible et d'interprétation délicate dans les coupes histologiques (Fig. ) , elle est formée par un canalicule excréteur soit simple, représentant sa totalité lorsqu’il demeure isolé jusqu’à la terminaison (glande simple) (Fig.), soit de 1er ordre lorsqu’il conflue ensuite (glandes ramifiée), dans les deux cas d’un même diamètre, subcylindrique et à paroi (épaisseur 600 Å) réduite à une seule couche, homogène, régulière, formée par de l’épicuticule. Les
canaux d’ordres supérieurs (2e ordre et excréteur principal) ont un diamètre plus grand (1,2 µ et 2 à 5 µ), une lumière plus large et une paroi formée cette fois par deux couches : l’une interne, épicuticulaire, dense, opaque et homogène comme celle du canal de 1er ordre, mais d’épaisseur irrégulière, paraissant festonnée en coupes transversales; l’autre externe, dérivée de l’exocuticule (?), plus claire, striée, montrant des fibres hélicoïdales et concentriques. Les canalicules excréteurs sont entourés par des cellules canalaires: une seule lorsque le canal est simple (Argyrodes cognatus) (Fig), plusieurs qui se succèdent en gaine continue lorsque la glande est ramifiée (Argyrodes argyrodes), dont une proximale, autour du canal de 2e ordre, et les autres distales, autour du conduit principal. Dans tous les cas l’adénocyte s’unit à la cellule canalaire adjacente par une jonction adhérente annulaire (zonula adherens). Peu visibles dans les coupes histologiques (pigmentation mélanique éventuelle, long noyau très basophile), les cellules canalaires montrent en M.E.T., qu’elles s’enroulent autour des conduits et renferment dans leur hyaloplasme clair des microtubules ainsi que quelques grains de pigment mélanique. Le noyau est très allongé, aplati, indenté, réniforme en coupe transversale et contient une chromatine abondante, dense et marginée. L’enroulement de chaque cellule détermine une invagination en méso de son plasmalemme (Fig.), avec jonction septée. D’autres jonctions l’unissent aux voisines.
Interprétation et rôles
Interprétation anatomique
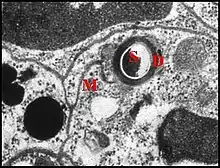
La structure fine des unités constituant la glande acronale permet de les rattacher à la classe 3 des cellules glandulaires épidermiques d’Arthropodes dans sa forme la plus simple : un canal est d’abord entouré par une seule cellule glandulaire où il débute, puis par une cellule canalaire et est enfin en continuité avec la cuticule au niveau de sa terminaison. Noirot et Quennedey (1974[10]) ont défini pour la première fois cette classe chez les insectes où existe également un appareil terminal (end apparatus), censé caractéristique de glandes exocrines tégumentaires défensives ou sécrétrices de phéromones. On sait par ailleurs que dans de nombreuses glandes exocrines d’insectes (salivaires, dermiques, annexes du tractus génital), l’appareil de Golgi ne paraît pas polarisé, ne se dispose pas en dictyosomes mais paraît uniquement formé par des vésicules. Il en est probablement de même pour la glande acronale des Argyrodes où les amas vésiculaires marqueraient l’emplacement de réseaux transgolgiens. Les unités peuvent être discrètes et isolées (types acuminé et nasuté) ou former un organe glandulaire anatomiquement défini, plus ou moins volumineux et massif (autres types) montrant généralement une bipartition visible dans les coupes histologiques transversales et la disposition symétrique de pores au M.E.B. La glande acronale d’Argyrodes fissifrontella (Seychelles) se présente comme une variation importante, pouvant exister aussi chez d’autres espèces, et qui a valeur de stade évolutif.
Interprétation fonctionnelle
La glande acronale est étroitement liée au comportement sexuel des Argyrodes .Les conformations étranges du prosoma des mâles ont été considérées comme « des bizarreries de la Nature … évidemment sans utilité » (Berland, 1932) et illustrées par Millot (1968 : fig. 479, p. 696) sans tentative d’explication. Il convenait de rechercher dans l’éthologie d’ Argyrodes certains traits de comportement pouvant relier des hypothèses fonctionnelles à l’existence de la glande. La relative immunité dont bénéficient les Argyrodes dans leurs rapports avec l’Araignée-hôte n’est nullement concernée. La glande acronale ne peut être considérée comme un organe de défense élaborant une substance répulsive ou vulnérante : elle manque chez les femelles ; l’absence de musculature compressive et la terminaison des canaux excréteurs dans une région anfractueuse conformée en cul de sac, n’impliquent pas une projection de substance.
De plus, dans le cas du type rostré, le seul, semble-t-il, dont l’accouplement et ses préludes aient été étudiés[6] - [2] jusqu'à la rédaction initiale de ce texte, les chélicères de la femelle étreignent la bosse frontale du mâle, leur doigt mobile pénétrant selon Legendre dans le repli - plus précisément l’échancrure- formé par la bosse frontale et le clypeus - plutôt la protubérance oculaire. Il est probable qu'au cours de cette manœuvre durant laquelle le mâle est littéralement prisonnier des chélicères…(Legendre)[2], les parties buccales de la femelle entrent en contact intime avec l’échancrure et la sécrétion clypéale qui doit s’y déverser. De nouvelles recherches anatomiques, axées cette fois sur le modelé des appendices péri-buccaux femelles, pourraient établir certaines correspondances entre ce dernier et les reliefs clypéaux mâles.
Il ne semble pas que la sécrétion soit consommée par la femelle (contrairement à ce qui se passe chez certains Insectes pour des glandes d'une autre nature : Dictyoptères et Panorpes) mais plutôt qu’elle s’évapore dans la dépression où les poils pourraient jouer un rôle pour la concentrer et (ou) la diffuser. Sa fonction est celle d’une phéromone qui facilite l’accouplement, donc aphrodisiaque, soit en inhibant momentanément l’agressivité de la femelle dont Legendre[2] a d’ailleurs souligné une « certaine torpeur» (effet pacificateur), soit en provoquant son excitation sexuelle (« arousal»[6]) ce qui paraît plus douteux.
En outre, et contrairement à une autre assertion de Whitehouse[6], rien ne prouve que la substance soit élaborée en permanence par le mâle qu’elle marquerait ainsi chimiquement.
Il est à noter que la copulation s’accompagne de la mise en place d’un bouchon d'accouplement (mating plug des auteurs de langue anglaise) sur l’épigyne femelle, sécrétion possible du pédipalpe mâle introduit. Chez des Araignées d'autres familles, ce bouchon pourrait être élaboré par les glandes gnathocoxales.
Annexes
Bibliographie
- C. Noirot et A. Quennedey, Fine structure of Insect epidermal glands, Ann.Rev.Entom., 19, 1974, p. 61-80[10].
Notes et références
- A. Lopez, avec R. Legendre, « Étude histologique de quelques formations glandulaires chez les Araignées du genre Argyrodes (Theridiidae) et description d’un nouveau type de glande : la glande clypéale des mâles », Bull. Soc. zool. Fr., 99 (3), p. 453-460.,
- Legendre,R., « Quelques remarques sur le comportement des Argyrodes malgaches (Araignées, Theridiidae). », Ann.Sci.nat., Zool., 12, p.507-512.,
- A. Lopez, « Contribution à l'étude des caractères sexuels somatiques chez les mâles des Aranéides », Thèse de Doctorat d'État es Sciences, Université de Montpellier II. 117 pp + planches (dont 4 en couleurs),
- A. Lopez, avec M. Emerit, « Données complémentaires sur la glande clypéale des Argyrodes (Araneae, Theridiidae). Utilisation du microscope électronique à balayage », Rev. Arachnol., 2 (4), p. 143-153, (lire en ligne)
- A. Lopez, avec M. Emerit, « The clypeal gland of Argyrodes fissifrontella Saaristo, 1978 (Araneae, Theridiidae). Bull. Br. Arachnol. Soc., 5 (4), p. 166-168 », Bull. Br. Arachnol. Soc., 5 (4), p. 166-168,
- M.E.A. Whitehouse, « The external structure detail of the protrusions on the cephalothorax of mâle Argyrodes antipodiana », Bull.Brit.Arachnol.Soc., 7, p. 142-144,
- A. Lopez, avec L. Juberthie-Jupeau et M. Emerit, « The « clypeal » gland of Argyrodes cognatus (Blackwall, 1877), a Theridiid Spider from the Seychelles Islands. Verhandl.8. Intern. Arachnol. Kongr., Wien 1980, p. 309-313 », Verhandl.8. Intern. Arachnol. Kongr., Wien 1980, p. 309-313,
- A. Lopez, avec R. Legendre, « Ultrastructure de la glande clypéale des mâles d’Araignées appartenant au genre Argyrodes (Theridiidae). », C.R.Acad.Sci. Paris, 281, p. 1101-1103,
- A. Lopez, avec C. Juberthie, « La glande clypéale d’Argyrodes argyrodes (Walck.) : nouvelles précisions sur son ultrastructure. », Rev.Arachnol., 3, p. 1-11,
- C. Noirot et A. Quennedey, « Fine structure of Insect epidermal glands. », Ann.Rev.Entom.,19, p. 61-80,