Entraînement des horloges périphériques chez les mammifères
L’horloge circadienne chez les mammifères est un système hiérarchisé d’oscillateurs générant un rythme de 24 h à partir de signaux lumineux du cycle jour-nuit de l’environnement[1]. En effet, les gènes impliqués dans les mécanismes de l’horloge sont présents non seulement dans le noyau suprachiasmatique (NSC), soit l’oscillateur principal, mais aussi dans les organes périphériques, soit les oscillateurs secondaires[2]. La persistance de l’expression de ces gènes de manière rythmique lorsque les tissus périphériques sont isolés du NSC démontre l’existence d’une horloge endogène omniprésente pour pratiquement toutes les cellules du corps[2]. Ce rythme circadien indépendant du NSC n’est cependant observé que pour quelques jours in vitro[3]. L’atténuation est probablement causée par une diminution progressive de l'amplitude de l'expression des gènes de l’horloge dans chaque cellule et une perte de synchronisation entre les cellules[4]. Par conséquent, le noyau suprachiasmatique n’est pas le générateur de l’horloge circadienne, mais il est plutôt considéré comme le chef d’orchestre[5].
De plus, l’activité physiologique circadienne, par exemple le rythme éveil-sommeil, est complètement altérée en présence de lésions au niveau de l’hypothalamus[6]. Ainsi, le noyau suprachiasmatique est essentiel à la synchronisation de la phase des horloges périphériques pour tous les tissus du corps qui eux, contrairement au NSC, ne sont pas sensibles à la lumière[4]. L’entraînement des horloges périphériques est notamment possible grâce à une combinaison de signaux neuronaux et endocriniens ainsi que via les fluctuations de la température corporelle et de l’horaire d’alimentation[1] - [7]. De plus, une altération du cycle local d’alimentation, d’hydratation[8] ou de température corporelle peut entrainer un changement de phase au niveau d’organes spécifiques en envoyant des signaux différents de ceux provenant du NSC[7]. Cet effet est prononcé dans le cas de l’alimentation puisqu’il peut affecter la phase des cycles de température corporelle, de régulation homéostatique et même de relâchement de glucocorticoïdes[7]. Comme la figure 1 le démontre, tous les cycles énoncés sont d’ailleurs des composantes non négligeables des horloges périphériques.
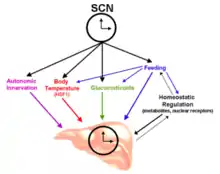
Signal d’entrée de l’horloge circadienne
Le noyau suprachiasmatique (NSC) se synchronise à l’environnement grâce à des photorécepteurs spécifiques, soit les cellules ganglionnaires rétiniennes intrinsèquement photosensibles (ipRGC) contenant de la mélanopsine, qui capte l’information lumineuse[9]. Celle-ci constitue le signal d’entrée dominant qui définit la phase du rythme circadien. Le signal est acheminé à l’oscillateur principal grâce à une cascade de signalisation via le tractus rétino-hypothalamique, qui active par la suite les gènes de l’horloge du NSC[5] - [10]. Ce dernier relaie ensuite l’information aux horloges secondaires situées dans les organes cibles via différentes voies décrites plus bas. Ces oscillateurs dits « esclaves » régulent ensuite à leur tour les rythmes locaux physiologiques et comportementaux[1]. En effet, les gènes circadiens présents dans les différents organes sont similaires, mais leur expression diffère selon chaque tissu spécifique afin de s’adapter de manière optimale à la fonction respective de ce tissu tout au long du cycle circadien[5]. Par conséquent, l’existence d’horloges secondaires dans les cellules périphériques tissulaires est utile pour l’optimisation de la physiologie cellulaire en isolant temporellement les réactions étant incompatibles chimiquement[4]. L’origine évolutive de ces processus provient probablement du fait que si ces différentes réactions étaient toutes gérées par l’horloge principale dans le cerveau, un très grand nombre de messagers spécifiques serait requis[4].
Voies d'entraînement des horloges périphériques
Signalisation directe
Par les voies des systèmes sympathique et parasympathique, le système nerveux autonome sert d’intermédiaire au niveau des voies de signalisation engendrées par le NSC ayant pour but l’entraînement des rythmes des horloges périphériques. Cette transmission semblerait être intimement liée à la sécrétion rythmique de glucocorticoïdes par l’entremise du système sympathique ainsi qu’au système de sécrétion rythmique de la corticolibérine (CRH) et de son successeur, l’hormone corticotrope (ACTH)[7]. L’ACTH serait aussi reliée à une régulation temporelle de la sensibilité et à l’amplitude de la réponse de sécrétion en glucocorticoïdes au niveau de l’horloge adrénale corticale. Elle serait donc en partie responsable de la régulation circadienne du relâchement des glucocorticoïdes[11]. L’action du système parasympathique est d’ailleurs responsable de la rythmicité de la sécrétion de la mélatonine au niveau de la glande pinéale. Cette hormone est liée à la rythmicité du cycle éveil-sommeil[12]. Les voies afférentes du système nerveux permettent donc au NSC de mieux synchroniser la phase des différents tissus périphériques grâce à l’intégration des signaux engendrés par les différents organes[13].
Perturbations circadiennes
Des études démontrent que certaines professions ayant des horaires instables ou de nuit, corrèlent avec des plus hautes prévalences de troubles métaboliques tels qu’une diminution de la capacité du système nerveux autonome à réguler les rythmes circadiens[14]. En conséquence, la rythmicité circadienne et son maintien sont affectés. De plus, l’horaire d’alimentation aurait un impact sur la régulation autonome du rythme cardiaque et du métabolisme des lipides, en affectant les composantes circadiennes au niveau de l'entraînement des horloges périphériques[15]. Ces découvertes supportent le lien existant entre les différents signaux entrants au niveau des horloges périphériques (c.-à-d. l' alimentation de nuit) et l’effet de changement de phase par rapport au cycle du NSC, ce dernier étant synchronisé à la lumière[14] - [15].
Rôle des glucocorticoïdes
L’utilisation en laboratoire d’un analogue de glucocorticoïdes a permis d’établir que ces derniers sont des agents entraînants au niveau des tissus périphériques in vivo[16]. Les glucocorticoïdes agissent probablement conjointement avec les éléments de réponse présents au niveau des gènes responsables de protéines cycliques. Celles-ci sont associées au rythme circadien de l’horloge centrale dans le NSC, et sont aussi présentes au niveau des tissus périphériques[7]. Cet effet de synchronisation dû aux glucocorticoïdes au niveau des horloges périphériques fut notamment observé au niveau des transcriptomes du foie chez la souris; la composante oscillatoire peut être rétablie chez des souris ayant subi des lésions au niveau du NSC dans l’hypothalamus grâce aux glucocorticoïdes. Ces derniers activeraient le récepteur nucléaire HNF4α[17] qui contient des boîtes-E pouvant interagir avec PER2 ainsi qu’au niveau du contrôle de la transcription des gènes CLOCK et BMAL1[18]. Ces gènes sont des composantes principales de l’horloge circadienne chez les mammifères, tant dans le NSC que dans les tissus et horloges périphériques. Plusieurs autres récepteurs nucléaires seraient aussi sensibles aux glucocorticoïdes de façon à synchroniser la transcription de gènes circadiens dans les tissus périphériques[7].
Désynchronisation du cycle jour-nuit par l’alimentation
L’expression des gènes de l’horloge circadienne dans les cellules périphériques est intimement liée à l’alimentation[19]. En effet, dans certains tissus périphériques, la nourriture serait le zeitgeber dominant des horloges périphériques chez plusieurs espèces de mammifères. L’imposition de régime alimentaire sous des conditions lumière-noirceur ou noirceur-noirceur peut changer la phase des rythmes circadiens de l’expression des gènes au niveau des cellules périphériques sans affecter la phase du NSC[19]. Puis, il est tout aussi possible de resynchroniser les horloges périphériques par des changements abrupts de l’horaire d’alimentation (régime alimentaire)[19]. Par exemple, le foie est l’organe qui est le plus sensible au régime alimentaire[20]. Effectivement, le foie joue un rôle dominant dans le métabolisme des glucides, des protéines et des lipides[19]. Il a été démontré qu’une partie du métabolisme du glucose (entre autres) est sous la régulation des rythmes circadiens[4] - [21]. De façon globale, ce phénomène pourrait être dû, entre autres, aux métabolites induits par la digestion de molécules comme le glucose et les hormones gastro-intestinales qui pourraient agir comme zeitgebers des horloges périphériques[4]. Toutefois, cet entraînement par la nourriture peut prendre quelques jours et peut être traité différemment selon le tissu[19].
Le Minh et al. ont établi que les glucocorticoïdes inhibent/ralentissent les changements de phases que pourrait induire l’imposition du régime alimentaire allant à l’encontre de la phase du NSC dans les horloges périphériques. Cependant, il est difficile de déterminer si cette inhibition par les glucocorticoïdes est induite par le NSC ou par le régime alimentaire lui-même[22].
De plus, la nourriture peut induire des changements drastiques de la température corporelle participant aussi à l'entraînement des horloges périphériques[19]. Donc, de façon générale, l’expression rythmique des gènes représente les voies de sortie des oscillateurs périphériques qui, en condition normale, sont synchronisés au NSC. Cependant, lorsqu’un régime alimentaire s’oppose à la phase du NSC sur une longue période, les oscillateurs périphériques deviennent désynchronisés du NSC. Puisque l’imposition d’un régime alimentaire n’affecte pas le NSC, ceci suppose qu’il entraîne indirectement les horloges périphériques par des cycles d’éveil-sommeil et donc d’alimentation[22].
Rôle de la température corporelle
Dans la majorité des organismes, la température est un agent important influençant les rythmes circadiens[7]. Par contre, les cycles de variation de température extérieure sont de faibles agents d’entraînement chez les mammifères[23]. Ceci est dû au fait que les homéothermes ont la capacité de maintenir leur température interne en dépit des fluctuations de la température ambiante[7]. La température corporelle des mammifères démontre néanmoins une oscillation circadienne régie par le NSC indépendamment du cycle éveil-sommeil[24], le NSC étant lui-même dépendant de la lumière pour la synchronisation de l’organisme avec le milieu extérieur[5]. De plus, les rythmes d’oscillation de la température corporelle sont aussi dépendants des habitudes de prise de nourriture et des cycles éveil-sommeil, étant eux-mêmes des sorties du NSC[4]. Les différents taux de réentraînement dans les tissus périphériques (contenant les oscillateurs périphériques) à un nouveau cycle lumière-noirceur suggèrent que ces derniers utilisent des combinaisons uniques de signaux pour se synchroniser au NSC[3]. Par contre, certains signaux semblent suffisants pour contrôler la phase de la majorité des tissus périphériques[5]. La température semble être un de ces signaux puisque les changements de température physiologique peuvent entrainer tous les oscillateurs périphériques ayant été examinés[25] - [26].
Le contrôle de la température interne est donc un signal de sortie du NSC agissant comme signal d’entrée sur les horloges circadiennes des tissus périphériques[5]. Plusieurs oscillateurs périphériques sont extrêmement sensibles aux changements de température, notamment les fibroblastes, le foie, les reins et les poumons[25] - [26] - [27] - [28]. Quant à lui, le NSC est résistant aux fluctuations physiologiques de la température corporelle[27]. Ce dernier n’étant pas influencé par le paramètre qu’il contrôle, les cycles de température circadiens représentent un signal d’entraînement idéal et universel pour les oscillateurs périphériques[7].
Le mécanisme d’action par lequel la température semble influencer les oscillateurs périphériques serait directement lié à l’interaction existant entre les Heat Shock Factors (HSF) et le gène mPer2[28]. Comme le démontre la figure 2, la température influencerait donc l’activité des HSF en régulant l’expression et la transcription de mPer2 au niveau local. Cette régulation se ferait probablement par l’entremise des Heat-Shock Elements (HSE), qui se retrouvent dans la région 5’ en amont du gène. Les HSF se lient aux HSE, ce qui induirait possiblement le changement de phase de l’horloge périphérique dû à la température[28].
Références
- Steven M. Reppert et David R. Weaver, « Coordination of circadian timing in mammals », Nature, vol. 418, no 6901, , p. 935–941 (ISSN 0028-0836, PMID 12198538, DOI 10.1038/nature00965, lire en ligne, consulté le )
- Seung-Hee Yoo, Shin Yamazaki, Phillip L. Lowrey et Kazuhiro Shimomura, « Period2:Luciferase real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues », Proceedings of the National Academy of Sciences of the United States of America, vol. 101, no 15, , p. 5339–5346 (ISSN 0027-8424, PMID 14963227, DOI 10.1073/pnas.0308709101, lire en ligne, consulté le )
- (en) Hajime Tei, Michael Menaker, Yoshiyuki Sakaki et Gene D. Block, « Resetting Central and Peripheral Circadian Oscillators in Transgenic Rats », Science, vol. 288, no 5466, , p. 682–685 (ISSN 1095-9203 et 0036-8075, PMID 10784453, DOI 10.1126/science.288.5466.682, lire en ligne, consulté le )
- Ueli Schibler, Juergen Ripperger et Steven A. Brown, « Peripheral circadian oscillators in mammals: time and food », Journal of Biological Rhythms, vol. 18, no 3, , p. 250–260 (ISSN 0748-7304, PMID 12828282, DOI 10.1177/0748730403018003007, lire en ligne, consulté le )
- Ethan D. Buhr et Joseph S. Takahashi, « Molecular components of the Mammalian circadian clock », Handbook of Experimental Pharmacology, no 217, , p. 3–27 (ISSN 0171-2004, PMID 23604473, PMCID PMC3762864, DOI 10.1007/978-3-642-25950-0_1, lire en ligne, consulté le )
- C. I. Eastman, R. E. Mistlberger et A. Rechtschaffen, « Suprachiasmatic nuclei lesions eliminate circadian temperature and sleep rhythms in the rat », Physiology & Behavior, vol. 32, no 3, , p. 357–368 (ISSN 0031-9384, PMID 6463124, lire en ligne, consulté le )
- Jennifer A. Mohawk, Carla B. Green et Joseph S. Takahashi, « Central and peripheral circadian clocks in mammals », Annual Review of Neuroscience, vol. 35, , p. 445–462 (ISSN 1545-4126, PMID 22483041, PMCID PMC3710582, DOI 10.1146/annurev-neuro-060909-153128, lire en ligne, consulté le )
- F. K. Stephan et I. Zucker, « Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions », Proceedings of the National Academy of Sciences of the United States of America, vol. 69, no 6, , p. 1583–1586 (ISSN 0027-8424, PMID 4556464, lire en ligne, consulté le )
- S. Hattar, H. W. Liao, M. Takao et D. M. Berson, « Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity », Science (New York, N.Y.), vol. 295, no 5557, , p. 1065–1070 (ISSN 1095-9203, PMID 11834834, PMCID PMC2885915, DOI 10.1126/science.1069609, lire en ligne, consulté le )
- R. Y. Moore et N. J. Lenn, « A retinohypothalamic projection in the rat », The Journal of Comparative Neurology, vol. 146, no 1, , p. 1–14 (ISSN 0021-9967, PMID 4116104, DOI 10.1002/cne.901460102, lire en ligne, consulté le )
- Henrik Oster, Sebastian Damerow, Silke Kiessling et Vladimira Jakubcakova, « The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock », Cell Metabolism, vol. 4, no 2, , p. 163–173 (ISSN 1550-4131, PMID 16890544, DOI 10.1016/j.cmet.2006.07.002, lire en ligne, consulté le )
- (en) « The suprachiasmatic nucleus controls the circadian rhythm of heart rate via the sympathetic nervous system », Physiology & Behavior, vol. 55, no 6, , p. 1091–1099 (ISSN 0031-9384, DOI 10.1016/0031-9384(94)90392-1, lire en ligne, consulté le )
- T. Ueyama, K. E. Krout, X. V. Nguyen et V. Karpitskiy, « Suprachiasmatic nucleus: a central autonomic clock », Nature Neuroscience, vol. 2, no 12, , p. 1051–1053 (ISSN 1097-6256, PMID 10570479, DOI 10.1038/15973, lire en ligne, consulté le )
- Surriya Jabeen, « Effect of Sleep/Wake Cycle on Autonomic Regulation », Journal of the College of Physicians and Surgeons--Pakistan: JCPSP, vol. 25, no 6, , p. 434–437 (ISSN 1681-7168, PMID 26100997, lire en ligne, consulté le )
- Takahiro Yoshizaki, Yuki Tada, Azumi Hida et Ayaka Sunami, « Effects of feeding schedule changes on the circadian phase of the cardiac autonomic nervous system and serum lipid levels », European Journal of Applied Physiology, vol. 113, no 10, , p. 2603–2611 (ISSN 1439-6327, PMID 23922171, DOI 10.1007/s00421-013-2702-z, lire en ligne, consulté le )
- A. Balsalobre, S. A. Brown, L. Marcacci et F. Tronche, « Resetting of circadian time in peripheral tissues by glucocorticoid signaling », Science (New York, N.Y.), vol. 289, no 5488, , p. 2344–2347 (ISSN 0036-8075, PMID 11009419, lire en ligne, consulté le )
- Akhilesh B. Reddy, Elizabeth S. Maywood, Natasha A. Karp et Verdun M. King, « Glucocorticoid signaling synchronizes the liver circadian transcriptome », Hepatology (Baltimore, Md.), vol. 45, no 6, , p. 1478–1488 (ISSN 0270-9139, PMID 17538967, DOI 10.1002/hep.21571, lire en ligne, consulté le )
- Isabelle Schmutz, Jürgen A. Ripperger, Stéphanie Baeriswyl-Aebischer et Urs Albrecht, « The mammalian clock component PERIOD2 coordinates circadian output by interaction with nuclear receptors », Genes & Development, vol. 24, no 4, , p. 345–357 (ISSN 1549-5477, PMID 20159955, PMCID PMC2816734, DOI 10.1101/gad.564110, lire en ligne, consulté le )
- Francesca Damiola, Nguyet Le Minh, Nicolas Preitner et Benoît Kornmann, « Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus », Genes & Development, vol. 14, no 23, , p. 2950–2961 (ISSN 0890-9369, PMID 11114885, lire en ligne, consulté le )
- (en) Yoshida C , et al., « Early nocturnal meal skipping alters the peripheral clock and increases lipogenesis in mice. - PubMed - NCBI », sur www.ncbi.nlm.nih.gov (consulté le )
- K. Ishikawa et T. Shimazu, « Daily rhythms of glycogen synthetase and phosphorylase activities in rat liver: influence of food and light », Life Sciences, vol. 19, no 12, , p. 1873–1878 (ISSN 0024-3205, PMID 826747, lire en ligne, consulté le )
- N. Le Minh, F. Damiola, F. Tronche et G. Schütz, « Glucocorticoid hormones inhibit food-induced phase-shifting of peripheral circadian oscillators », The EMBO journal, vol. 20, no 24, , p. 7128–7136 (ISSN 0261-4189, PMID 11742989, DOI 10.1093/emboj/20.24.7128, lire en ligne, consulté le )
- R. Refinetti et M. Menaker, « The circadian rhythm of body temperature », Physiology & Behavior, vol. 51, no 3, , p. 613–637 (ISSN 0031-9384, PMID 1523238, lire en ligne, consulté le )
- (en) « Environmental light and suprachiasmatic nucleus interact in the regulation of body temperature », Neuroscience, vol. 132, no 2, , p. 465–477 (ISSN 0306-4522, DOI 10.1016/j.neuroscience.2004.12.012, lire en ligne, consulté le )
- (en) Joseph S. Takahashi, Seung-Hee Yoo et Ethan D. Buhr, « Temperature as a Universal Resetting Cue for Mammalian Circadian Oscillators », Science, vol. 330, no 6002, , p. 379–385 (ISSN 1095-9203 et 0036-8075, PMID 20947768, DOI 10.1126/science.1195262, lire en ligne, consulté le )
- (en) « Rhythms of Mammalian Body Temperature Can Sustain Peripheral Circadian Clocks », Current Biology, vol. 12, no 18, , p. 1574–1583 (ISSN 0960-9822, DOI 10.1016/S0960-9822(02)01145-4, lire en ligne, consulté le )
- (en) Hanspeter Herzel, Achim Kramer, Markus Heine et Pål O. Westermark, « Coupling governs entrainment range of circadian clocks », Molecular Systems Biology, vol. 6, no 1, , p. 438 (ISSN 1744-4292 et 1744-4292, PMID 21119632, DOI 10.1038/msb.2010.92, lire en ligne, consulté le )
- (en) Ueli Schibler, Joseph S. Takahashi, Hermann Bujard et Olivier Schaad, « System-Driven and Oscillator-Dependent Circadian Transcription in Mice with a Conditionally Active Liver Clock », PLOS Biology, vol. 5, no 2, , e34 (ISSN 1545-7885, PMID 17298173, PMCID PMC1783671, DOI 10.1371/journal.pbio.0050034, lire en ligne, consulté le )