Dientamoeba fragilis
Dientamoeba fragilis est une espèce de parasite, retrouvée dans le tractus gastro-intestinal de plusieurs espèces dont l'humain, le cochon ou le gorille. Il est composé de différents génotypes non distinguables par microscopie conventionnelle, mais pouvant présenter une pathogénicité différente (source : L. Gétaz F. Chappuis L. Loutan, Parasitoses intestinales et hépatiques : diagnostic et traitement Rev Med Suisse 2007; volume 3. 32314) la cause de troubles digestifs[2]. Il est une cause fréquente de diarrhées du voyageur, diarrhées chroniques, asthénie et, chez l'enfant, de troubles du développement.

| Domaine | Eukaryota |
|---|---|
| Règne | Excavata |
| Embranchement | Metamonada |
| Classe | Parabasalia |
| Ordre | Trichomonadida |
| Famille | Monocercomonadidae |
| Genre | Dientamoeba |
Étymologie
- Di se réfère aux deux noyaux visibles chez le trophozoïte (stade durant lequel le parasite se nourrit).
- Ent fait référence à l'environnement intestinal dans lequel le micro-organisme est retrouvé.
- Le nom d'espèce fragilis se réfère au fait que les trophozoïtes soient fragiles. En effet, il ne survivent pas longtemps dans les selles après l'excrétion.
Il a été décrit pour la première fois en 1918 par M. W. Jepps et C. Dobell[1] - [3].
Dientamœbose
L'infection à D. fragilis, appelée dientamœbose, est associée de manière variable aux symptômes suivants : douleurs abdominales, diarrhée, perte de poids, fièvre. Dans une étude, D. fragilis a été observé chez 0,9% des patients observés. Sa co-incidence avec l'oxyurose a été rapportée[4].
Biologie
Dientamoeba fragilis est un parasite protozoaire appartenant au phylum des Parabasalia, proche du genre Trichomonas. Il fait donc partie des protozoaires flagellés bien qu’il en soit dépourvu [5]. Il a été décrit pour la première fois en 1918 par M. W. Jepps et C. Dobell[1] et était alors considéré comme une amibe binucléées ayant une faible résistance en dehors de son hôte, ces éléments justifiant l’appellation ayant toujours cours aujourd’hui[1]. Il s’agit d’un micro-organisme très communément retrouvé dans les selles de sujets humains, sains comme symptomatiques, de distribution mondiale. Il semble cependant exister peu d’hôtes non-humains à ce protozoaire qui n’a sinon été décrit que chez certains primates non-humains (gorilles, macaques, babouins)[6] - [7] ou récemment chez le cochon[8]. Ainsi, on ne décrit pour l’instant que 2 génotypes de D. fragilis, le génotype 1 étant largement plus fréquent que le génotype 2 (dit souche Bi/PA)[5]. Cependant, l’étude de la diversité génétique de D. fragilis n’est pas très avancée, peu d’équipes travaillant sur ces questions, et se basant principalement sur l’étude des séquences du gène de la sous-unité 18S de l’ARN ribosomique. Certains auteurs suggèrent cependant que la pathogénicité variable du micro-organisme pourrait être due à une variabilité génétique plus grande qui pourrait être explorée par l’étude génétique de gènes de ménage autres que les séquences d’ARN ribosomique[8].
Morphologie
Le trophozoïte est caractérisé par la présence de deux noyaux la plupart du temps, bien qu’un seul puisse être identifié dans 20 à 40 % des cas[1]. Le cytoplasme contient en général une vacuole et peut contenir des débris issus de l’alimentation, en forme de granulations. Il peut cependant apparaître uniforme. La taille du micro-organisme peut varier de manière importante, allant de 5 à 15 μm de diamètre, et la forme globale de la cellule peut être arrondie ou ovalaire[5]. Comme indiqué précédemment, malgré son caractère génétique de flagellé, D. fragilis ne possède pas de flagelle et se déplace à la manière d’une amibe par émission de pseudopodes cytoplasmiques[6]. Il existe dans la forme bi-nucléée une structure en tige entre les deux noyaux servant de point de départ à certaines formations polaires adjacentes à un des noyaux. Ces formations s'avèrent être analogues aux actratophores de protozoaires hypermastigotes. On retrouve également un appareil de Golgi. La structure nucléaire de D. fragilis est plus proche de celle des flagellés que celle des organismes du genre Entamoeba. La présence d'hydrogénosomes est également notable, étant caractéristique d'autres trichomonades[9].
Le kyste est de description beaucoup plus récente puisqu’il a été identifié une première fois en 2013 sur des modèles murins puis en 2014 dans des échantillons de selles humaines, les deux fois par la même équipe[10] - [11]. De manière assez surprenante, l’étude par microscopie électronique de ce kyste retrouve des organites spécifiques à la présence d’un flagelle, telles que l’axostyle ou des axonèmes flagellaires, sans qu’il n’y ait effectivement un flagelle extériorisé. Ces éléments sont cohérents avec le caractère phylogénétique de flagellé de D. fragilis, et il a été proposé par les auteurs ayant fait ces observations que le protozoaire se serait adapté à la vie entérique en perdant son flagelle afin d’adopter un mode de déplacement amiboïde[11]. Par ailleurs, ce kyste présente une paroi épaisse, qualifiée de « mur », et une structure nucléaire similaire à celle du trophozoïte.
Enfin, il a été décrit un stade de pré-kyste, caractérisé par une forme compacte, sphérique et homogène, plus petite que le trophozoïte (4 à 5 μm). Ce stade n’a été décrit qu’une seule fois, par une seule équipe de recherche[10] - [5].
Cycle de vie
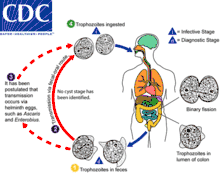
Le cycle de D. fragilis fait probablement intervenir les différentes formes précédemment décrites. L’existence d’une forme kystique rendrait fort probable une transmission manuportée féco-orale, puisque représentant une forme de résistance permettant au parasite de survivre dans le milieu extérieur ce qui n’est pas le cas pour le trophozoïte. Cependant, le caractère récent des descriptions des formes kystique et pré-kystique ne permet pas encore d’affirmer que cette voie de contamination est possible. Avant la découverte de ces formes, il avait été postulé que la transmission soit médiée par des œufs d’helminthes telles qu’Enterobius vermicularis ou Ascaris lumbricoides. Cette hypothèse se fondait sur la proximité entre D. fragilis et le protozoaire Histomonas meleagridis, parasite de la poule dont la transmission inter-individuelle est assurée via les œufs d’Heterakis gallinae, nématode parasite des gallinacés[12]. Des études génétiques avaient été effectuées, à la recherche d’ADN de D. fragilis à la surface d’œufs d’E. vermicularis dont les résultats avaient été positifs[13]. Les deux hypothèses ne sont, en tout état de cause, pas antinomiques, une transmission de D. fragilis médiée par des œufs d’helminthes étant envisageable, que le protozoaire soit sous forme kystique ou de trophozoïte. Le reste du cycle se déroule dans le tube digestif de l’hôte où le protozoaire, sous forme trophozoïte se multiplie par fission binaire et se nourrit de bactéries de la faune digestive par phagocytose.
Références
- M. W. Jepps, C. Dobell, « Dientamoeba fragilis ng, n. sp., a new intestinal amoeba from man. », Parasitology, vol. 10, , p. 352-367 (DOI 10.1017/S0031182000003929, lire en ligne)
- Windsor JJ, Macfarlane L, « Irritable bowel syndrome: the need to exclude Dientamoeba fragilis », Am. J. Trop. Med. Hyg., vol. 72, no 5, , p. 501; author reply 501–2 (PMID 15891119, lire en ligne)
- Johnson EH, Windsor JJ, Clark CG, « Emerging from obscurity: biological, clinical, and diagnostic aspects of Dientamoeba fragilis », Clin. Microbiol. Rev., vol. 17, no 3, , p. 553–70, table of contents (PMID 15258093, PMCID 452553, DOI 10.1128/CMR.17.3.553-570.2004, lire en ligne)
- Stark D, Beebe N, Marriott D, Ellis J, Harkness J, « Prospective study of the prevalence, genotyping, and clinical relevance of Dientamoeba fragilis infections in an Australian population », J. Clin. Microbiol., vol. 43, no 6, , p. 2718–23 (PMID 15956388, PMCID 1151954, DOI 10.1128/JCM.43.6.2718-2723.2005, lire en ligne)
- Garcia LS, « Dientamoeba fragilis, one of the neglected intestinal protozoa. », Journal of Clinical Microbiology, (PMID 27053676)
- Windsor JJ, Johnson EH, « Dientamoeba fragilis: the unflagellated human flagellate. », British journal of biomedial science, vol. 56, no 4, , p. 293 - 306 (PMID 10795375)
- Stark D, Phillips O, Peckett D, Munro U, Marriott D, Harkness J, Ellis J, « Gorillas are a host for Dientamoeba fragilis: an update on the life cycle and host distribution. », Veterinary parasitology, vol. 151, no 1, , p. 21-6 (PMID 18022187, lire en ligne)
- Cacciò SM, Sannella AR, Manuali E, Tosini F, Sensi M, Crotti D, Pozio E, « Pigs as natural hosts of Dientamoeba fragilis genotypes found in humans. », Emerging infectious disease, vol. 18, no 5, , p. 838-841 (PMID 22515838, lire en ligne)
- Tachezy, Jan: Hydrogenosomes and mitosomes: mitochondria of anaerobic eukaryotes (ISBN 978-3-642-09542-9)
- Stark D, Garcia LS, Barratt JL, Phillips O, Roberts T, Marriott D, Harkness J, Ellis JT, « Description of Dientamoeba fragilis cyst and precystic forms from human samples. », Journal of clinial microbiology, vol. 52, no 7, , p. 2680-3 (PMID 24808242, lire en ligne)
- Munasinghe VS, Vella NG, Ellis JT, Windsor PA, Stark D, « Cyst formation and faecal-oral transmission of Dientamoeba fragilis--the missing link in the life cycle of an emerging pathogen. », International journal for parasitology, vol. 43, no 11, , p. 879-83 (PMID 23872523, lire en ligne)
- Clark CG, Röser D, Stensvold CR, « Transmission of Dientamoeba fragilis: pinworm or cysts? », Trends in parasitology, vol. 30, no 3, , p. 136-40 (PMID 24492020, lire en ligne)
- Ögren J, Dienus O, Löfgren S, Iveroth P, Matussek A, « Dientamoeba fragilis DNA detection in Enterobius vermicularis eggs. », Pathogens and disease, vol. 69, no 2, , p. 157-8 (PMID 23893951, lire en ligne)