Canalisation (biologie)
La canalisation est, en biologie du développement, un modèle théorique et expérimental proposé par Wagner (en). Il s'agit de la mesure de la capacité d'un groupe d'individus à conserver le même phénotype malgré la variabilité de son environnement ou de son génotype[1].
Historique et concepts
La première approche de ce concept fut apportée par Waddington (1942) disant que pour un caractère bien adapté – et donc proche de l’optimum – tout agent diminuant les effets néfastes des mutations devrait être sélectionné. Par la suite, dans les années 1990, il devint beaucoup plus facile d’expérimenter sur les gènes grâce aux techniques moléculaires. Cela comprend également les multiples expérimentations sur le modèle de la Drosophile. Finalement, toutes ces expérimentions démontrant les multiples variations phénotypiques possibles convergent pour dire – comme M. Andreas Wagner lui-même – qu’il existe une sorte de « tampon évolutionnaire génétique » résistant aux diverses perturbations environnementales et/ou génétiques (aussi définies comme « interactions épigénétiques » selon M. Wagner). Autrement dit, « des environnements stressants peuvent pousser à l’expression de variations génétiques cachées »[2]. Selon Wagner (1996), ce phénomène pourrait s’expliquer par le fait que des pressions de sélection longuement exercées sur des gènes et régulateurs transcriptionnels – hautement sollicités dans les patrons de développement de l’individu – auraient pour effet de diminuer leur taux de mutations. C’est ici que des termes comme « stabilité épigénétique » ou « canalisation développementale » prennent leur sens[3] - [4].
Dans tous les cas, l’on parle ici de l’interaction entre gènes et l'environnement ou encore de l’effet des environnements sur les gènes. Il est donc important de comprendre ce que signifie et comprend la notion d'interactions entre gènes – unités de base héréditaires donnant un caractère – et environnement(s), et ce dans un contexte développemental d’organismes multicellulaires. Cela signifie que ces interactions doivent passer au travers une hiérarchie de différents niveaux d’organisation : « le gène; le cytoplasme; la cellule; les organes ; les systèmes composés d’organes; l’individu (et ses comportements) qui lui est soumis aux forces de l’environnement qui lui sont externes »[5].
Aujourd’hui, plusieurs autres termes synonymes peuvent être utilisés pour décrire le concept de canalisation tels que : les traits canalisés, allèles homéorhétiques ou encore un tampon développemental. La canalisation s’oppose par principe à la notion de « plasticité développementale » (ou plus simplement « plasticité phénotypique ») étant « la capacité d'un organisme à exprimer différents phénotypes à partir d'un génotype donné selon des conditions environnementales ». Donc ces deux concepts opposés ont un rôle évolutif dans le sens de (1) « permettre - au cours du développement ou pas - l’accumulation de variations génétiques » et (2) « être des mécanismes donnant au phénotype une sensibilité ou insensibilité face à l’environnement »[6]).
Puisque la canalisation peut impliquer des gènes ayant un rôle développemental, il est bon d'introduire la notion des homéogènes pour simplifier la complexité du développement. Un gène dit homéotique est un gène tenu responsable du développement et possédant une homéoboîte. Cette dernière est une séquence propre aux homéogènes comprenant 180 nucléotides codant une protéine possédant à son tour un homéodomaine, donc une homéoprotéine. Comme pour les homéogènes, l’homéodomaine – motif (ou portion) conservé de 60 acides aminés permettant la liaison à l’ADN – est propre aux homéoprotéines. Ces homéogènes ont différents rôles lors de différents stades développementaux, contribuant pour résumer à produire « les bonnes parties du corps au bon endroit ». Le plus souvent cela peut inclure des signaux émis de leur part vers des tissus cibles : des morphogènes. Encore une fois, la canalisation peut intervenir pour limiter les changements et les variations possibles vers un phénotype stable au cours du développement (ou pas, si l’organisme est déjà pleinement formé et mature). Donc, le phénotype résulte alors de l’interaction de ces deux extrêmes que sont les signaux géniques et environnementaux (épigénétiques)[7].
Exemples actuels chez des organismes modèles

La Drosophile est un insecte de l’ordre des Diptères ; elle est plus connue sous le nom de « mouche à vinaigre/fruits ». Elle comporte des centaines d’espèces et est très sollicitée comme organisme modèle, car elle présente un organisme à développement rapide et incluant un passage par le stade larvaire. Ceci explique en partie les multiples expériences exercée sur cette mouche concernant l’altération du phénotype sauvage via le développement. Du point de vue génétique, elle possède quatre paires de chromosomes constituant l’intégralité de son matériel génétique (ou génome) aujourd’hui séquencé. Plusieurs aspects de la canalisation peuvent être observés via des expérimentations[8].
Héritabilité de la canalisation
L'une des premières expériences effectuées fut celle de Waddington et Rendel sur divers gènes pour la plupart homéotiques dont le gène « scute ». Le gène scute est pro-neuronal car il est impliqué dans la neurogenèse (formation des gènes) en plus d’être un responsable de la détermination des sexes et le développement des poils scutellaires (poils sur une partie thoracique). Ces poils scutellaires sont normalement en nombre de quatre chez les drosophiles. Rendel effectua des mutations pour diminuer le nombre de ces poils (de 0 à 3) et effectua des sélections artificielles (élevage sélectif des animaux). Il vit par la suite à l’analyse des générations suivantes que la majorité des organismes avaient encore quatre poils scutellaires. X’était donc ici un phénotype canalisé puisqu’il était le plus répandu et qu’il fallait accumuler beaucoup de mutations pour en différer. Cependant, Rendel alla même jusqu’à faire muter leur processus de canalisation lui-même et démontra que la canalisation est héritable. En fait, il créa des groupes de drosophiles à deux poils scutellaires et vit que les générations suivantes conservaient ce même nombre de poils. Cet exemple ci-dessus est démonstratif de l'effet de la canalisateur sur un gène en particulier mais aussi de son héritabilité[1] - [9].
Canalisation et patron de développement
La canalisation peut également agir sur les réseaux de gènes, dont ceux sollicités lors du développement via les voies de signalisation. Une voie de signalisation est une voie de transduction de signal cellulaire permettant aux cellules de communiquer entre elles via des signaux moléculaire interagissant avec des protéines. La voie de signalisation BMP (Bone Morphogenetic Protein, un ligand) permet la spécification des cellules dorsales en plus du bon développement des amnioserosa. Les amnioserosa sont des cellules épithéliales remplissant les cavités épidermiques dorsales à un stade embryobiologie)|nnaire de la drosophile appelé gastrulation (formation des axes embryonnaires). Donc ici le bon fonctionnement de cette voie BMP - impliquée dans le passage du stade embryonnaire blastula à gastrula - est assuré par un réseau génique composé de trois gènes : egr, cv-2 et zen. Chacun de ces gènes a un rôle bien particulier. Egr (eiger) induit l’augmentation du signal étant la synthèse de BMP (ou rétroaction positive) ; Cv-2 (crossveinless-2) induit l’inhibition ; et Zen (zerknüllt) est une homéoboîte assurant précocement la transcription des deux premiers gènes pour qu’ils puissent agir[10] - [11] - [12].
Dans une expérience de Galvin-Smyth et al. sur ce système développemental décrit ci-haut, il fut démontré qu’en l’absence (1) des gènes egr et cv-2, il y a une sérieuse augmentation/variation dans la signalisation de BMP mais pas quand l’un ou l’autre des deux gènes est présent ; (2) la même observation s’appliqua en l’absence de zen. Donc, cela démontre que le bon signalement de la voie BMP est un processus canalisé bistable car il dépend des deux étapes ou composantes génétiques différentes : l’expression de zen puis celle de egr et cv-2 (ou l’un des deux), et ce dans le but de permettre l’expression appropriée des cellules amnioserosa au bon endroit (partie dorsale de l’embryon) et bon moment (gastrulation). Donc dans ce contexte, l‘expression des cellules amnioserosa est le phénotype canalisé. Un autre aspect intéressant est que c’est un phénotype canalisé à plusieurs niveaux car il peut encore y avoir bon développement (spécification) des cellules amnioserosa même en cas de baisse du signal de BMP et la variation dans le nombre de cellules n’affecte pas la viabilité de l’embryon car ce dernier y démontre une tolérance. Tout ceci est démonstratif de l’effet de la canalisation au niveau des réseaux de gènes; mais ça ne s’arrête pas là.
De façon subséquente, il est même démontré que la canalisation peut agir dans d’autres perspectives par exemple de façon interspécifique : de façon différente pour différentes espèces d’un même genre/groupe. Après avoir compris le fonctionnement de la voie BMP, Galvin-Smyth et al. commencèrent par chercher la cause de l’expression précoce du gène zen. Ils trouvèrent en partie qu’elle est initiée par un « élément répresseur ventral » conservé en amont par rapport à zen et contenant des sites de liaisons permettant la liaison d’un facteur transcriptionnel de type « doigt de zinc » nommé Zelda. Pour que ce dernier puisse bien se lier à cet élément régulateur répresseur il doit avoir des motifs (les sites de liaison) de type « TAG » ; sans eux – comme avec des motifs mutés – le facteur Zelda ne peut pas se lier et il s’ensuit une diminution de l’expression de zen. Donc cela représente un autre élément sous l’effet ou le contrôle de la canalisation sur les gènes pour le bon développement dorso-ventral de la drosophile.
Cependant, deux autres aspects additionnels sont remarquables dans ce contexte. D’abord, en comparant différents les stades embryonnaires développementaux, on peut observer des conséquences différentes au sujet de l’expression de zen ou egr (succédant à zen). Par exemple, il a été observé que des mutations sur ces motifs TAG atténuaient l’expression au stade 5e embryonnaire mais pas au 6e. Cela signifie que la canalisation peut aussi agir qu’à partir de certaines périodes spécifiques lors du développement, et cela a été visible pour toutes les espèces de drosophiles utilisées. Ensuite, Galvin-Smyth et al. les trois espèces de Drosophila utilisée sont : D. melanogaster, D. yakuba et D. santomea. Bien que ces trois espèces proviennent de lieux différents, l’effet de l’expression atténuée du gène zen aboutit ultimement à des conséquences différentes quant au nombre de cellules amnioserosa. Effectivement, pour le même stade embryonnaire chez les trois espèces il a été observé que l’espèce D. santomea avait le plus haut taux de variation comparativement aux deux autres espèces de Drosophila. Cela démontre l’effet interspécifique de la canalisation : elle induit un effet stabilisation chez beaucoup d’individu d’un même groupe (comme D. melanogaster et D. yakuba) mais pas pour certains (ici D. santomea). En bref, ici l’on voir que l’espèce D. santomea est beaucoup plus susceptible aux mutations : dans ce contexte l’on peut parler d’espèce ‘’décanalisée’’.
Les quatre paragraphes ci-haut représentent un bel exemple de toute la complexité d’application de la canalisation : son action peut être définie à partir d’une certaine période, sur certaines espèces, avec la présence de certains éléments, etc. Comme l’on peut le voir, même si la canalisation tend à la stabilité des individus elle peut avoir des ‘’champs d’action’’ où elle ne peut pas vraiment agir. L’exemple suivant, également assez complexe en termes d’éléments et conséquences impliqués, démontre bien ce dernier point : les limites que peut avoir la canalisation[10].
Frontière entre canalisation et stabilité développementale chez la souris

Il existe également des exemples expérimentaux de canalisation très poussée chez la souris. La souris (ou Mus musculus) est un autre organisme modèle très sollicité en laboratoire. Cet un eucaryote mammifère appartenant à l’ordre des rongeurs. Comme pour la Drosophila melanogaster, elle a un cycle de reproduction rapide, mais a plus de paires chromosomiques (20) par rapport à la Drosophile en plus de son génome séquencé. Étant un organisme plus complexe, l’impact de la canalisation peut impliquer plus d’effets variables à divers niveaux génétiques comme morphologiques. L’expérience d’Hallgrimsson B. et al. est un bon exemple de l’impact d’un gène muté sur la variation phénotypique lors du développement du crâne de la souris. Il est possible de voir que la canalisation a une certaine limite dans la restriction de la fluctuation des phénotypes et que cela peut empiéter sur un autre processus stabilisateur[13] - [14].
Pour décrire tous les éléments de ce système et ce qu’il importe, commençons par le gène Papss2. Ce dernier code la phosphoadénosine-phosphosulfate synthétase 2 (une protéine) en plus d’être aussi impliqué dans la bonne croissance/disposition du cartilage crânien (il n’est donc pas forcément un homéogène). Le rôle de cette protéine – chez la souris comme chez l’homme – est d’induire une sulfatation (réaction biochimique impliquant le ‘’transfert de groupes sulfates’’) sur une autre molécule : c’est une sulfotransférase (ou une enzyme). Cette dernière cible diverses molécules biologiques incluant des protéines comme les glycosaminoglycanes (aussi nommées ‘’GAGs’’). Pour que Papss2 puisse effectuer sa fonction elle doit être sous une forme fonctionnelle/active/phosphorylée assurée par le gène Papss2 lui-même. Si la combinaison fonctionnelle des allèles – par exemple les génotypes A/A comme A/a fonctionnent mais pas a/a – est mutée cela peut induire de graves impacts sur la formation crânienne de l’individu, comme c’est le cas si le gène a pour forme (bm/bm). Cela nous permet d’introduire la mutation « Brachyomorphe »[15].
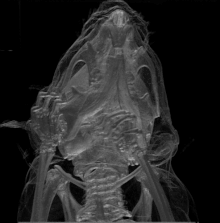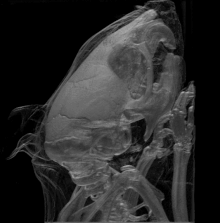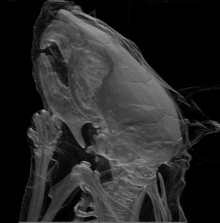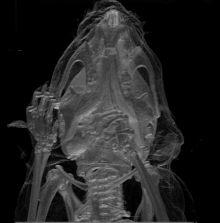
La mutation Brachyomorphe est une mutation autosomique récessive (mutation au niveau des chromosomes corporels; non sexuels) causée par la substitution d’une paire de bases. Elle résulte d’un changement d’acide aminé au niveau de l’enzyme (sulfotransférase) décrite ci-haut. L’impact de gène muté finit ultimement par atténuer la croissance du cartilage via la composition de sa matrice extracellulaire. En fait, le gène muté ne peut plus assurer la forme active de l’enzyme (via sa phosphorylation); cette dernière ne peut pas sulfater les glycosaminoglycanes devenant donc moins accessibles via agrégation et donc altérant ainsi la composition de la matrice extracellulaire. C’est cette dernière conséquence dans qui finit par réduire la croissance du cartilage et tous les éléments osseux en dépendant. Au niveau crânien, il est possible de voir que différentes zones anatomiques en sont affectées en termes de taille et forme (se référer à l’image 4) : le neurocrâne; la base du crâne (ou plancher de la cavité crânienne); temporal/zygomatique faisant référence aux joues/pommettes et côtés du crâne; le visage (au niveau antérieur du crâne).
Les expériences d’Hallgrimsson B. et al. ont réalisé en testant des groupes standardisés (groupe contrôle) et expérimentaux qu’il y a avait une tendance similaire quant aux fluctuations individuelles au sein des 2 groupes et ce au niveau de la taille et forme des squelettes. Cette covariation (variation dans la forme et la taille) était bien sûr plus évidente dans le cas du groupe expérimental mais démontre que la canalisation n’est plus en mesure d’agir à ce niveau de complexité via la présence de la fluctuation asymétrique constance (déviation de ce qui représente une parfaite symétrie bilatérale à cause de plusieurs facteurs stressants)[16].
Le fait de parler de fluctuation asymétrie constante – pour des mutants tous homozygotes ou génétiquement pareils – fait référence aux « déviations individuelles autour d’une moyenne/norme » et on parle donc du concept de stabilité développementale, et non pas de canalisation. Pour que la canalisation eût agi ici, il aurait fallu voir des variations uniquement dans le groupe expérimental avec de très faibles variations phénotypiques (dans la forme ou/et la taille du crâne) dans le groupe contrôle (devant conserver le phénotype sauvage). Cela serait tout à fait typique de ce qui serait attendu sous l’effet de la canalisation agissant sur la stabilité dans la variation phénotypique. La raison exacte de la non-application de la canalisation dans cet exemple est encore inconnue, mais en se référant à l’exemple précédent (sur la drosophile) et en regardant toute la complexité de ce système développemental il est bien plausible que certains « éléments » manquent pour qu’elle puisse agir[13].
Canalisation, stabilité développementale et plasticité
Cette dernière parenthèse sert d’ouverture pour illustrer que la canalisation, la stabilité développementale et la plasticité ont toutes leur mécanisme propre et agissent toutes pour permettre aux individus de se rapprocher d’un ‘’optimum’’ (défini selon l’environnement ou le génotype). Ces trois concepts - principalement la canalisation dans notre cas – ont probablement tous joué un rôle évolutif pour aboutir à la diversité de la vie observée aujourd’hui[17]
Références
- Meiklejohn, C.D., and Hartl, D.L. (2002). A single mode of canalization. Trends in Ecology et Evolution 17, 468–473.
- (en) Hermisson, J., and Wagner, G.P. (2004). The Population Genetic Theory of Hidden Variation and Genetic Robustness. Genetics 168, 2271–2284.
- (en) Wagner, A. (1996). Does Evolutionary Plasticity Evolve? Evolution 50, 1008–1023.
- Huerta-Sanchez, E., and Durrett, R. (2007). Wagner’s canalization model. Theoretical Population Biology 71, 121–130.
- (en) Gottlieb, G. (1991). Experiential canalization of behavioral development: Theory. Developmental Psychology 27, 4–13.
- « Plasticité et Canalisation », sur lbbe.univ-lyon1.fr (consulté le ).
- « De Homéoboîte à Homéoprotéine », sur bio-top.net (consulté le ).
- « Drosophile-mouche du vinaigre », sur Futura Sciences (consulté le ).
- (en) « Interactive Fly, Drosophilia », sur sdbonline.org (consulté le ).
- (en) Gavin-Smyth, J., Wang, Y.-C., Butler, I., and Ferguson, E.L. (2013). A Genetic Network Conferring Canalization to a Bistable Patterning System in Drosophila. Current Biology 23, 2296–2302.
- « Gene Ontology Term: amnioserosa formation ».
- « Compréhension Neuve de la façon dont le panneau de cellules entaille », News Medical Life Sciences, (lire en ligne).
- (en) Hallgrímsson, B., Brown, J.J.Y., Ford-Hutchinson, A.F., Sheets, H.D., Zelditch, M.L., and Jirik, F.R. (2006). The brachymorph mouse and the developmental-genetic basis for canalization and morphological integration. Evolution et Development 8, 61–73.
- « Souris de laboratoire », sur Futura Sciences (consulté le ).
- (en) « PAPSS2 3'-phosphoadenosine 5'-phosphosulfate synthase 2 [ Homo sapiens (human) ] », National Center for Biotechnology Information (consulté le ).
- (en) « Fluctuating Asymmetry », sur animalbehavioronline.com (consulté le ).
- (en) Vincent Debat et Patrice David, « Mapping phenotypes: canalization, plasticity and developmental stability », Trends in Ecologyand Evolution, vol. 16, no 10, , p. 555-561 (lire en ligne).