Activation CRISPR
L'activation CRISPR (CRISPRa) est un type d'outil CRISPR qui utilise des versions modifiées des effecteurs CRISPR sans activité endonucléase, avec des activateurs transcriptionnels ajoutés sur dCas9 ou les ARN guides (ARNg)[1].
Comme pour l'interférence CRISPR, l'effecteur CRISPR est guidé vers la cible par un guide ARN complémentaire (ARNg). Cependant, les systèmes d'activation CRISPR sont fusionnés à des activateurs transcriptionnels pour augmenter l'expression des gènes d'intérêt. De tels systèmes sont utilisables à de nombreuses fins, tels que les criblages génétiques et la surexpression de protéines d'intérêt. L'effecteur le plus couramment utilisé est basé sur Cas9, mais d'autres comme Cas12a ont également été utilisés[2].
Composants
dCas9
Cas9 avec endonucléase inactive (dead Cas9 ou dCas9), est une forme mutante de Cas9 dont l'activité endonucléase est supprimée par des mutations ponctuelles dans ses domaines endonucléases. Semblable à sa forme non mutée, dCas9 est utilisé dans les systèmes CRISPR avec des ARNg pour cibler des gènes ou des nucléotides spécifiques complémentaires de l'ARNg avec des séquences PAM (motif de reconnaissance du proto-espaceur (en)) qui permettent à Cas9 de se lier. Cas9 possède généralement 2 domaines d'endonucléase appelés domaines RuvC et HNH. Les mutations ponctuelles D10A et H840A modifient 2 résidus importants pour l'activité de l'endonucléase qui aboutissent finalement à sa désactivation. Bien que dCas9 n'ait pas d'activité endonucléase, il est toujours capable de se lier à son ARN guide et au brin d'ADN ciblé, car cette liaison est gérée par d'autres domaines. Cela suffit souvent à atténuer, voire à bloquer la transcription du gène ciblé si l'ARNg positionne dCas9 d'une manière qui empêche les facteurs transcriptionnels et l'ARN polymérase d'accéder à l'ADN. Cependant, cette capacité à se lier à l'ADN peut également être exploitée pour l'activation puisque dCas9 possède des régions modifiables, généralement les extrémités N et C de la protéine, qui peuvent être utilisées pour attacher des activateurs transcriptionnels[3].
ARN guide
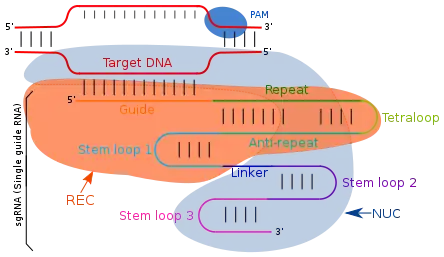
Un petit ARN guide (sgRNA pour small guide RNA, ou gRNA an anglais) est un ARN d'environ 20 nucléotides utilisé pour diriger Cas9 ou dCas9 vers leurs cibles. Les ARNg contiennent deux régions majeures importantes pour les systèmes CRISPR : les régions d'échafaudage et d'espacement. La région espaceur a des nucléotides qui sont complémentaires de ceux trouvés sur les gènes cibles, souvent dans la région promotrice. La région d'échafaudage est responsable de la formation d'un complexe avec (d)Cas9. Ensemble, ils lient (d)Cas9 et le dirigent vers le(s) gène(s) d'intérêt. Étant donné que la région d'espacement d'un ARNg peut être modifiée pour n'importe quelle séquence potentielle, ils donnent aux systèmes CRISPR beaucoup plus de flexibilité car tous les gènes et nucléotides avec une séquence complémentaire à la région d'espacement peuvent devenir des cibles possibles[3].
Activateurs transcriptionnels
Les activateurs transcriptionnels sont des domaines protéiques ou des protéines entières liées à dCas9 ou à des sgRNA qui aident au recrutement de cofacteurs importants ainsi qu'à l'ARN polymérase pour la transcription du ou des gènes ciblés par le système. Pour qu'une protéine soit fabriquée à partir du gène qui la code, l'ARN polymérase doit produire de l'ARN à partir de la matrice d'ADN du gène au cours d'un processus appelé transcription. Les activateurs transcriptionnels ont un domaine de liaison à l'ADN et un domaine pour l'activation de la transcription. Le domaine d'activation peut recruter des facteurs de transcription généraux ou une ARN polymérase dans la séquence du gène. Les domaines d'activation peuvent également fonctionner en facilitant la transcription par des ARN polymérases bloquées et, chez les eucaryotes, peuvent agir pour déplacer les nucléosomes sur l'ADN ou modifier les histones pour augmenter l'expression des gènes[4]. Ces activateurs peuvent être introduits dans le système par fixation à dCas9 ou au sgRNA. Certains chercheurs ont noté que l'étendue de la régulation positive de la transcription peut être modulée en utilisant plusieurs sites pour la fixation de l'activateur dans une expérience et en utilisant différentes variations et combinaisons d'activateurs à la fois dans une expérience ou un échantillon donné[5] - [6] - [7].
Système d'expression
Un système d'expression est nécessaire pour l'introduction des protéines gRNAs et (d)Cas9 dans les cellules d'intérêt. Les options typiquement employées comprennent les plasmides et les vecteurs viraux tels que ceux des virus adéno-associé (AAV) ou lentivirus.
Systèmes d'activation spécifiques
VP64-p65-Rta
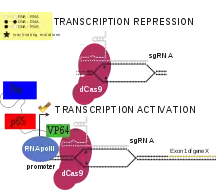
L'activateur VP64-p65-Rta, ou VPR, dCas9 a été créé en modifiant un activateur dCas9 existant, dans lequel un activateur transcriptionnel Vp64 est joint à l'extrémité C-terminale de dCas9[1]. Dans la protéine dCas9-VPR, les facteurs de transcription p65 et Rta sont ajoutés à l'extrémité C-terminale de dCas9-Vp64. Par conséquent, les trois facteurs de transcription sont ciblés sur le même gène. L'utilisation de trois facteurs de transcription, par opposition à Vp64 uniquement, entraîne une expression accrue des gènes ciblés. Lorsque différents gènes ont été ciblés par dCas9, ils ont tous montré une expression significativement plus élevée avec dCas9-VPR qu'avec dCas9-VP64. dCas9-VPR peut être utilisé pour augmenter l'expression de plusieurs gènes dans la même cellule en mettant plusieurs sgARN dans la même cellule[8]. dCas9-VPR a été utilisé pour activer les gènes de la neurogénine 2 et de la différenciation neurogène 1, entraînant la différenciation des cellules souches pluripotentes induites et d'astrocytes en neurones induits[8]. Une étude comparant les activateurs dCas9 a révélé que les activateurs VPR, SAM et Suntag fonctionnaient mieux avec dCas9 pour augmenter l'expression des gènes dans une variété de types de cellules de drosophiles, de souris et humaines.
Médiateur d'activation synergique
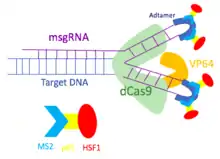
Pour surmonter la limitation du système d'activation du gène dCas9-VP64, le système dCas9-SAM a été développé pour incorporer plusieurs facteurs transcriptionnels. Utilisant les protéines MS2, p65 et HSF1, le système dCas9-SAM recrute divers facteurs transcriptionnels travaillant en synergie pour activer le gène d'intérêt. Afin d'assembler différents activateurs transcriptionnels, le système dCas9-SAM utilise un ARN guide unique modifié (sgRNA) qui possède des sites de liaison pour la protéine MS2. Les aptamères en épingle à cheveux sont attachés à la boucle tétra et à la tige-boucle 2 de l'ARNsg pour devenir des sites de liaison pour les protéines d'enveloppe dimérisées du bactériophage MS2. Comme les aptamères en épingles à cheveux sont exposées à l'extérieur du complexe dCas9-sgRNA, d'autres facteurs transcriptionnels peuvent se lier à la protéine MS2 sans perturber le complexe dCas9-sgRNA. Ainsi, la protéine MS2 est conçue pour inclure les protéines p65 et HSF1. La protéine de fusion MS2-p65-HSF1 interagit avec le dCas9-VP64 pour recruter plus de facteurs de transcription sur le promoteur des gènes cibles.
Applications
Le système d'activation dCas9 permet d'exprimer un gène souhaité ou plusieurs gènes dans la même cellule. Il est possible d'étudier les gènes impliqués dans un certain processus en utilisant un écran large du génome qui implique l'activation de l'expression des gènes. L'examen des sgARN qui produisent un phénotype suggère quels gènes sont impliqués dans une voie spécifique. Le système d'activation dCas9 peut être utilisé pour contrôler exactement quelles cellules sont activées et à quel moment l'activation se produit. Des constructions dCas9 ont été fabriquées qui activent une protéine de fusion dCas9-activateur en présence de lumière ou de produits chimiques. Les cellules peuvent également être reprogrammées ou différenciées d'un type cellulaire à un autre en augmentant l'expression de certains gènes importants pour la formation ou le maintien d'un type cellulaire[9].
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « CRISPR activation » (voir la liste des auteurs).
- (en) « Targeted regulation of transcription in primary cells using CRISPRa and CRISPRi », Genome Res, vol. 31, no 11, , p. 2120–2130 (PMID 34407984, DOI 10.1101/gr.275607.121).
- « Multiplexed orthogonal genome editing and transcriptional activation by Cas12a », Nat Methods, vol. 16, no 1, , p. 51–54 (PMID 30559432, DOI 10.1038/s41592-018-0262-1, S2CID 56174507).
- « CRISPR/Cas9 Guide », Addgene.
- Ma, J. (August 2011). Transcriptional activators and activation mechanisms. Protein and Cell, 2(11), 879-888.
- « RNA-guided gene activation by CRISPR-Cas9-based transcription factors », Nature Methods, vol. 10, no 10, , p. 973–6 (PMID 23892895, PMCID 3911785, DOI 10.1038/nmeth.2600).
- « CRISPR RNA-guided activation of endogenous human genes », Nature Methods, vol. 10, no 10, , p. 977–9 (PMID 23892898, PMCID 3794058, DOI 10.1038/nmeth.2598).
- « Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex », Nature, vol. 517, no 7536, , p. 583–8 (PMID 25494202, PMCID 4420636, DOI 10.1038/nature14136, Bibcode 2015Natur.517..583K).
- « Highly efficient Cas9-mediated transcriptional programming », Nature Methods, vol. 12, no 4, , p. 326–8 (PMID 25730490, PMCID 4393883, DOI 10.1038/nmeth.3312).
- « Beyond editing: repurposing CRISPR-Cas9 for precision genome regulation and interrogation », Nature Reviews. Molecular Cell Biology, vol. 17, no 1, , p. 5–15 (PMID 26670017, PMCID 4922510, DOI 10.1038/nrm.2015.2).