Évolution insulaire
L’évolution insulaire caractérise la modification des fréquences alléliques de populations isolées sur des îles représentant un domaine géographique sur lequel les échanges extérieurs sont limités, au cours des générations[1].

Les îles se forment de deux manières. Premièrement, elles peuvent surgir de la mer, ce sont des îles océaniques qui se forment au-dessus d’un point chaud. Elles sont donc vides de toute forme de vie au départ, et sont colonisées progressivement par les espèces du continent voisin ou des îles voisines. Deuxièmement, elles peuvent être d’origine continentale, résultant de la fragmentation du continent ou de la montée des eaux. Ces îles sont habitées par la faune et la flore du continent d’origine[1] - [2]. La fragmentation des milieux naturels causée par l’être humain, par exemple lors de construction d’autoroutes, conduit à la formation d'îles ou îlots de végétation. La notion d'île en écologie ne se cantonne pas uniquement aux îles au sens géographique, mais prend en compte tous types de territoires de petites tailles et isolés[1].
Histoire
Les milieux insulaires passionnent les écologues depuis plusieurs siècles, car les écosystèmes qu’ils abritent diffèrent de ceux observés sur le continent. L’étude de l’insularité a permis l'émergence de nombreuses théories sur l’évolution des espèces, la plus célèbre étant la théorie de l’évolution par la sélection naturelle de Darwin, qu’il a notamment illustré avec l’évolution de la taille du bec chez les pinsons des Galapagos. Les îles sont caractérisées par leur isolement, et représentent donc un espace plus contrôlé et plus propice à l’élaboration de modèles simples pour comprendre les mécanismes de l’évolution[1].
La théorie des équilibres dynamiques de MacArthur et Wilson (1967) sur la biogéographie insulaire a permis de comprendre les mécanismes qui sous-tendent les processus de colonisation, d’extinction et de renouvellement d’espèces sur une échelle de temps relativement courte. En revanche, ce modèle ne prend en compte que des facteurs géographiques, tels que la taille et l’isolement de l’île, et ne tient pas rigueur des processus évolutifs tels que la coévolution et la spéciation, qui se manifestent sur une échelle de temps plus longue[1].
Aujourd'hui, il y a d’importants enjeux de conservation à propos des espèces insulaires. En effet, de nombreuses espèces insulaires sont en danger d'extinction, d’autres sont totalement éteintes. Les causes sont multiples, de plus en plus associées aux activités anthropiques comme la fragmentation des habitats, la pollution ou l’introduction d’espèces invasives[1].
Mécanismes de l'évolution insulaire
Il existe quatre pressions évolutives, également appelées forces de l’évolution, qui peuvent modifier la structure génétique des populations. Chacune d’entre elles a des effets différents sur la structure génétique et la divergence des populations. Ces quatre forces évolutives sont le flux de gènes, la dérive génétique, la sélection naturelle et la mutation. Ces quatre forces évolutives agissent simultanément sur les populations.
Flux de gènes
Le flux de gènes se définit par le mouvement d’individus ou de propagules. Le mouvement des individus se caractérise par leur déplacement, que ce soit par le départ ou l’arrivée des individus dans une population. Cela permet de diminuer la différenciation génétique, c’est-à-dire les différences génétiques entre différentes populations[2].
Les îles sont caractérisées par leur isolement géographique, par conséquent, les flux d’individus, et donc de leurs gènes, sont limités, voire absents, entre les îles et les continents, ou même entre les îles. De plus, le flux de gènes et d’individus provoquant une homogénéité entre les populations, les espèces vivant sur les îles se différencient peu à peu de celles vivant sur le continent ou de l'île d’où elles proviennent.
Cette absence de flux de gènes entraîne une diminution de la diversité spécifique (ou nombre d'espèces différentes sur l'île)[3]. Ainsi, prédateurs et espèces compétitrices sont moins nombreux, ce qui influe sur l'évolution des espèces insulaires.
Dérive génétique
La dérive génétique est un phénomène ayant lieu dans toutes les populations mais ses effets sont d’autant plus forts que les populations sont de petite taille. Le flux de gènes entre les individus sur le continent et les individus colonisateurs étant réduit ou absent, la colonisation d’îles par une espèce est une situation où les populations ont peu d’individus sur l’île. La dérive génétique provoque une diminution de la diversité au sein des populations et une augmentation de la variation entre des populations différentes. Ainsi, sur une île, elle provoque une diminution de la diversité au sein de la population de colonisateurs mais une augmentation des différences entre les populations du continent et celles de l’île[2].
Sachant que dans les petites populations il y a une dominance de la dérive génétique par rapport à la sélection naturelle, les mutations délétères ont autant de probabilité d’être fixées dans la population que les mutations neutres et bénéfiques. Ceci contribue à la perte de diversité génétique et de la capacité d’adaptation, ce qui implique un gros risque d'extinction pour les petites populations en milieu insulaire.
Sélection naturelle
La sélection naturelle est la variation non aléatoire des fréquences d’allèles. Ce mécanisme est responsable de l’adaptation des organismes à leur environnement. Les conditions environnementales et géographiques des îles sont souvent différentes de celles des continents, ce qui conduit à des pressions de sélection différentes entre ces deux milieux. La sélection naturelle est donc impliquée dans les syndromes d’insularité et les autres phénomènes détaillés par la suite[1].
Mutation
Les mutations sont des événements aléatoires de modification d’un nucléotide dans la séquence génétique d’un individu. Le résultat de cette mutation peut générer un nouvel allèle si la mutation a lieu sur la séquence codante d’un gène. Par ailleurs, si cette mutation se situe sur un chromosome contenu dans les gamètes, elle peut être transmise à la descendance et se propager au sein de la population sur plusieurs générations, sous l’effet de la sélection naturelle ou de la dérive génétique[1]. Ainsi, les mutations augmentent la variabilité génétique, sont à l’origine des variations des traits phénotypiques et ont des effets aléatoires sur ces traits. Il y a donc autant de probabilité de voir apparaître de nouvelles mutations au sein des populations insulaires qu’au sein des populations continentales. En revanche, l’influence des mutations sur la structure génétique des populations est très faible, voire négligeable, par rapport aux autres pressions évolutives.
Spéciation insulaire
La spéciation est le phénomène qui conduit une espèce ancestrale à se différencier en deux espèces dérivées. On considère généralement que des espèces sont différentes quand elles sont isolées génétiquement et ne peuvent plus se reproduire entre elles[4].
La spéciation sur une île peut correspondre à la distinction entre une espèce sur l’île et une autre sur le continent mais aussi à la formation de plusieurs espèces différentes à partir d’une initiale sur l’île. La spéciation n’est pas un phénomène qui a obligatoirement lieu sur une île. Cependant, une spéciation semble s’opérer sur de nombreuses îles et taxons résultant de mécanismes de l’évolution, du hasard et de la divergence des lignées depuis des sources continentales[2].
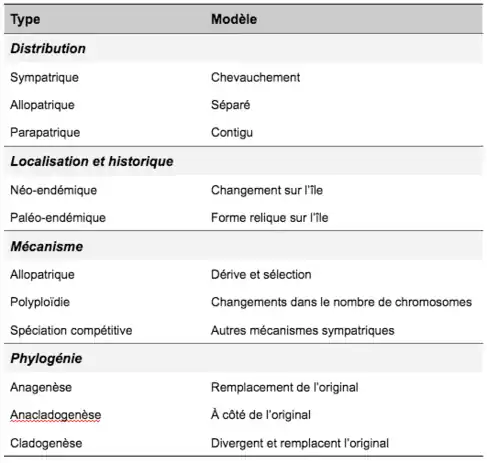
La spéciation peut avoir lieu dans le cas de colonisation d’une île mais elle peut aussi avoir lieu en dehors de ce processus. Plus généralement, la spéciation ayant lieu sur les îles a été étudiée de plusieurs manières (voir Tableau I)[4].
La spéciation insulaire peut s’envisager d’un point de vue historique et de localisation. Une espèce endémique est naturellement restreinte à un territoire précis. Pour les espèces endémique des îles, on fait une distinction entre :
- Des formes néo-endémiques qui ont évoluées in situ sur l’île
- Des formes paléo-endémique qui sont considérées comme endémiques par l’extinction des formes proches sur le continent. Elles peuvent alors être considérées comme des formes reliques.
En pratique, on ne peut pas toujours utiliser la comparaison des espèces sur le continent et sur l’île. En effet, les individus sur les continents issus de la même population que celles qui a fourni les individus colonisateurs ou qui se sont retrouvés sur une île lors de son isolement du continent, ont aussi évolué donc nous n’avons que rarement accès à cette population d’origine.
La spéciation insulaire peut s’envisager d’un point de vue géographique, et dans ce cas, nous pouvons distinguer :
- La spéciation allopatrique : les populations sont isolées géographiquement les unes des autres, soit entre l’île et le continent, soit entre îles ou encore au sein même d’une île. Toutes les espèces néo endémiques en sont un exemple.
- La spéciation sympatrique : les populations ne sont pas séparées géographiquement sur l’île. Lorsque les îles sont proches les unes des autres et que les organismes impliqués ont de grandes capacités de dispersion, il peut être difficile de distinguer la spéciation sympatrique de la spéciation allopatrique.
- La spéciation parapatrique : une partie des populations reste en contact entre elles mais la majorité des individus sont suffisamment éloignés pour ne pas se rencontrer.
- La spéciation dans la cas d’archipels est parfois utilisée pour caractériser spécifiquement les spéciations ayant lieu entre îles et peut impliquer à la fois de la sympatrie et de l’allopatrie.
En pratique, il est difficile de savoir quel type de spéciation a lieu. On considère généralement que le phénomène de spéciation allopatrique est dominant.
La spéciation insulaire peut s’envisager d’un point de vue des mécanismes avec différents processus :
- La spéciation géographique ou allopatrique étant la formation d’une barrière géographique limitant les flux de gènes dans la population reproductrice entre l’île et le continent. Ce phénomène à lieu à la fois dans le cas d’une colonisation d’île ou de la formation d’île se séparant du continent. Les différentes populations sont isolées et évoluent séparément. Elles deviennent assez différentes pour être considérées comme deux espèces différentes. Parfois, la barrière géographique se brise rendant possible des phénomènes de double invasion.
- La spéciation compétitive décrit un ensemble de modes de spéciation sympatrique. Cette spéciation est liée à la capacité d’individus d’une population présente sur l’île à étendre sa propre niche écologique en exploitant une niche écologique inoccupée. Les individus ayant exploité la nouvelle niche écologique et ceux restant dans la niche d'origine forment deux espèces différentes issues de la même population. Ce phénomène est possible sur une île car il y a moins de compétition avec les autres espèces et de niche occupée. La spéciation se produit notamment à cause d’une pression de compétition forte entre les deux populations.
- La polyploïdie caractérise l’augmentation du nombre de chromosomes. Ce phénomène peut mener à de la spéciation par deux processus : l’autopolyploidie c'est-à-dire la multiplication du génome d’une seule espèce parentale et l’allopolyploidie c’est-à-dire multiplication du génome des deux espèces parentales. Cette spéciation semble avoir particulièrement lieu pour des espèces de plantes sur les îles mais les raisons sont méconnues et ce résultat est controversé.
La spéciation insulaire peut finalement se considérer d’un point de vue de la structure de la lignée avec une distinction entre :
- L'anagenèse qui est la situation où l’espèce à l’origine de l'espèce actuelle est éteinte. Cela peut se produire au sein d’une île ou entre l’île et le continent.
- La cladogenèse, qui est la situation où deux espèces actuelles forment deux lignées issues de l’espèce d'origine qui s’éteint. Cela peut se produire sur l’île ou entre l’île et le continent.
- L’anacladogenèse, qui est la situation où l’espèce d’origine survit avec des petites divergences avec l’espèce descendante. Cela peut se produire sur la même île ou dans le même archipel.
La distinction entre les manières de caractériser la spéciation sur les îles peuvent ainsi se résumer dans le Tableau I :
Évolution lors de la colonisation des îles
La colonisation d’une île est le cas où des individus colonisateurs atteignent une île et forment une ou plusieurs populations fondatrices. Pour une île qui émerge de la mer, toute espèce présente sur l’île résulte d’un événement de colonisation. Ce n’est pas le cas pour les îles formées par la séparation avec un continent, qui présentent à la fois des espèces issues d’événements de colonisation, mais aussi des espèces qui étaient présentes sur l’espace qui s’est séparé du continent. Les populations fondatrices issues de l'événement de fondation peuvent être considérées comme biaisées car elles ne contiennent qu’une partie de la diversité génétique présente sur le continent. Elles sont aussi isolées car elles ne reçoivent plus ou très peu d’individus et de gènes du continent[2].
Lors de l’immigration et la colonisation d’une île, les meilleurs immigrateurs et les meilleurs colonisateurs sont sélectionnés[1] - [3]. Les effets de la sélection des individus qui immigrent le mieux peuvent durer longtemps après la colonisation, si les pressions de sélection liées à la colonisation ne vont pas à l’encontre des biais initiaux des populations fondatrices. Pour les individus immigrateurs, et notamment les vertébrés, les traits et capacités liés à l’immigration augmentent avec la taille corporelle. Les individus de plus grande taille devraient donc être favorisés au début de la colonisation (voir partie sur l’évolution de la taille)[3].
Effet fondateur
L’effet fondateur est la formation d’une nouvelle population à partir d’un très petit nombre d’individus. Il s’agit donc d’un phénomène ayant lieu dans des populations fondatrices dans le cas de la colonisation d’îles. Différents mécanismes évolutifs agissent sur ces populations biaisées et isolées[2] :
- Des mutations qui leur sont propres et peuvent augmenter la diversité génétique de ces populations insulaires et des différences par rapport aux populations des continents
- De la recombinaison comme la dérive génétique ayant un effet fort dans les petites populations et permettant la fixation d’allèles différents de la population d’origine sur le continent
- De la sélection naturelle produisant la sélection d’allèles particulièrement adaptés à l’environnement de l’île
Cependant la contribution de chacun de ces mécanismes à la spéciation est sujet à débat. Suivant les modèles, la colonisation et l’effet fondateur peuvent donner des effets différents pouvant conduire à la spéciation. Voici les 3 principaux modèles d’effet fondateur[2] - [5] :
- La spéciation péripatrique de Mayr : il s’agit de la spéciation de populations périphériques facilitée par l’évolution rapide des petites populations dans le cas d’un effet pionnier. La formation d’isolats périphériques et l’évolution indépendante des populations permettent la formation d’une nouvelle espèce. De plus, la petite taille de la population fondée va faire augmenter le coefficient de consanguinité au cours des premières générations. Ainsi, la probabilité que ces individus se retrouvent avec deux allèles récessifs et désavantageux qui ne s'expriment pas chez les hétérozygotes augmente. Ces allèles sont alors souvent éliminés par la sélection naturelle ce qui provoque une diminution de l’hétérozygotie dans ces populations. Les effets les plus importants dans ce modèle sont donc un manque d'hétérozygotie combiné avec de la dérive génétique.
- La spéciation fondatrice ou théorie des cycles d’expansion-catastrophe de Carson : Les effets les plus importants dans ce modèle sont le rôle des variations de pression de sélection et plus spécifiquement juste après la colonisation.
- La transilience génétique de Templeton : Les effets les plus importants dans ce modèle sont les changements dans quelques locus majeurs dans le génome.
Goulot d'étranglement
Un goulot d’étranglement est une réduction drastique et rapide de l’effectif d’une population. La perte d’un grand nombre d’individus et de leurs gènes provoque une baisse de diversité génétique dans la population après l'événement. En ce sens, la colonisation d’une île peut aussi être considérée comme un goulot d’étranglement[2]. On peut distinguer deux situations :
- Ce goulot d’étranglement peut être répété et fort, par exemple si la population colonisatrice initiale se disperse sur l’île et forment plusieurs populations colonisatrices isolées entre elles ou encore dans le cas de la colonisation d’archipels. Dans ce cas, la diversité génétique diminue à chaque séparation entre une population mère et une population fondatrice et ces effets fondateurs répétés produisent une diminution de l’hétérozygotie.
- Ce goulot d’étranglement peut avoir lieu une seule fois avec une population colonisatrice qui ne disperse pas et qui retrouve une grande taille rapidement. Dans ce cas, le goulot d’étranglement peut provoquer l’augmentation de la variance génétique en brisant la base génétique de certains complexes adaptatifs plus vieux. De nouveaux phénotypes peuvent en résulter dans les générations suivant le goulot d’étranglement, notamment s’ils sont soumis à l’action de la sélection naturelle.
Sélection sexuelle et introgression
La colonisation des îles a aussi un fort impact sur la sélection sexuelle. En effet, dans les populations de petites tailles, il y a sélection des femelles les moins discriminantes dans leur choix de mâles car le nombre de ces derniers est réduit. Cela augmente la possibilité de reproduction avec des populations d’espèces ou variétés proches qui peuvent aussi être présentes sur l’île. Des phénomènes d’hybridation ont donc lieu provoquant une augmentation de la diversité génétique dans la population fondatrice : c’est ce que l’on appelle l’introgression. L’importance de cet événement dans le processus de colonisation d’île a longtemps été sous-estimée avant que l’on accède aux méthodes d’études moléculaires[2].
Conclusion
La colonisation d’une île provoque donc la formation de populations de très petites tailles que l’on peut qualifier d’événement de fondation et de goulot d’étranglement et qui peuvent avoir différents effets[2] :
- La perte de diversité génétique et d’hétérozygotie à cause de l’effet fondateur et des goulots d’étranglements répétés
- La formation de nouvelles combinaisons génétiques sous l’effet d’un goulot d’étranglement unique
- La formation de variation génétique liée à l’hybridation entre des espèces ou variétés proches.
Ces effets provoquent une phase d’évolution rapide des populations présentes sur l’île. En général, la variabilité génétique entre les populations d’un continent et d’une île[2] :
- Diminue quand la taille des populations fondatrices augmente,
- Augmente quand la différence de taille entre la population sur le continent et sur l’île augmente,
- Diminue quand les capacités d’immigration et de dispersion des populations fondatrices augmentent,
- Diminue quand la taille de l’île augmente,
- Augmente quand la distance avec le continent augmente.
Syndrome d'insularité et manifestations
Les espèces installées en milieu insulaire subissent différents changements morphologiques, physiologiques, démographiques et comportementaux, que l’on appelle syndrome d'insularité[1] - [6].
Évolution de la taille
En milieu insulaire, de nombreux taxons, dont des plantes et des insectes, présentent des tailles très différentes de celles qu’ils ont sur le continent[2]. L’évolution peut causer des modifications de la taille des espèces par rapport à leurs ancêtres des continents, notamment chez les vertébrés où on constate des cas de gigantisme (les espèces de petite taille deviennent plus grandes sur les îles) et de nanisme (les espèces de grande taille deviennent plus petites sur les îles). La loi de Foster décrit ces modifications comme une tendance graduelle à travers et au sein des taxons, allant du gigantisme chez les plus petites espèces, au nanisme chez les espèces plus grandes[3] - [6] .
Causes
Cette évolution des tailles est principalement causée par la limitation en ressources, la diminution de la compétition interspécifique et la diminution de prédateurs. Plus une île est de petite taille et isolée du continent, plus la diversité spécifique, c’est-à-dire le nombre d’espèces et leur abondance relative, diminue. Il y a une plus grande diversité spécifique sur les surfaces les plus grandes, et donc plus d’espèces différentes sur les continents que sur les îles. Au contraire, la densité au sein de chaque espèce augmente (densité intraspécifique) sur les îles plus petites. On peut ainsi observer des densités similaires ou même plus élevées sur les îles que sur les continents. Les ressources sont moins abondantes sur une île ce qui conduit à une compétition intraspécifique (au sein d’une même espèce) pour les ressources plus intense, d’autant plus que la densité intraspécifique est aussi plus grande. Il y a donc une modification des forces de pression qui opèrent sur les îles : des interactions interspécifiques (entre différentes espèces) qu’on observe sur le continent (prédation, parasitisme, compétition) vers de la compétition intraspécifique. La diminution des pressions interspécifiques est un relâchement écologique (ecological release en anglais). Le relâchement écologique et la limitation des ressources sont les deux forces sélectives principales qui s’exercent sur populations insulaires[3].
Le relâchement écologique joue un rôle à la fois dans le gigantisme et le nanisme. Dans le cas du gigantisme, la diminution des plus grands prédateurs et compétiteurs conduit à une augmentation de la taille car ils n’ont plus besoin de se cacher ou de s’échapper. Pour les espèces dont les ancêtres continentaux étaient de grande taille afin de pouvoir se défendre contre les grands prédateurs, ce moyen de défense n’est plus utile étant donné la diminution des prédateurs. Le gigantisme est aussi favorisé par la sélection des individus qui immigrent, alors que le nanisme résulte de la limitation des ressources, il est plus difficile pour un organisme de grande taille de trouver suffisamment de nourriture[3].
Les causes du gigantisme et du nanisme ne font pas consensus parmi les chercheurs. Pour certains, le gigantisme et le nanisme sont dus à la limitation en ressource, qui induit de la compétition intraspécifique. Des espèces grandissent pour mieux faire face à la compétition tandis que d’autres vont évoluer vers une plus petite taille en réponse à cette diminution de ressources. La compétition intraspécifique est la cause principale de la sélection directionnelle d’une grande taille selon d’autres spécialistes. Enfin, ce serait la taille des îles qui déterminerait le facteur qui dirige la sélection. Sur les îles de petite taille, la limitation en ressource est responsable de la sélection alors que sur les îles de grande taille, il s’agit du relâchement écologique[6].
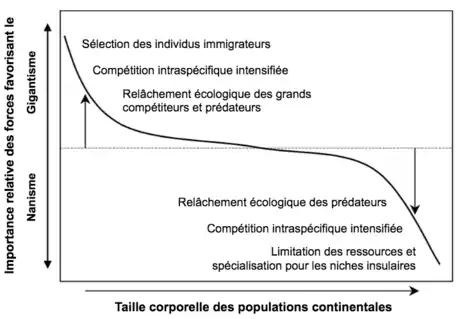
La loi de Foster résulte de la sélection naturelle d’individus et de populations en milieu insulaire, il y a une sélection directionnelle de certaines tailles selon les espèces. La sélection naturelle est donc le mécanisme responsable de l’évolution vers du gigantisme ou du nanisme[3] - [6].
Exemple de gigantisme et nanisme
La loi de Foster s’applique surtout chez les mammifères mais aussi chez les reptiles et les oiseaux[3].
Dans le cas du gigantisme, on peut citer :
- Les tortues géantes des Galapagos et de l’Océan Indien[2]
- Des espèces de rongeurs et de canards[2]
Pour le nanisme :
- Le paresseux des îles Bocas del Torro au Panama[6]
- Le mammouth laineux de l’île Wrangel, éteint[2] - [3]
- Les éléphants nains, aujourd’hui éteints, de Crète, Malte (Elephas falconeri), Sicile[2]
- Le crocodile nain Crocodylus niloticus en Mauritanie[2]
- Homo floresiensis, hominidé éteint de l’île de Flores en Indonésie, probablement issu de Homo erectus[2]
- Plusieurs espèces de lézard de petites îles des Caraïbes dont un lézard endémique de Sainte-Croix, Pholidoscelis polops[2]
- Des lignées dinosauriennes, telles Magyarosaurus, Telmatosaurus ou Zalmoxes dans l'île Maastrichtienne de Hatseg, située en mer Téthysienne, aujourd'hui devenue une dépression des Carpates, en Europe centrale[7]
Docilité insulaire
Le comportement des espèces insulaires est impacté par l’isolement des îles et évolue différemment de celui des espèces continentales. Le comportement, au même titre qu’un trait physique ou physiologique, est un phénotype dont on peut étudier l'évolution.
De manière générale, les individus insulaires semblent pour la plupart moins effrayés, voire plus dociles, que les espèces continentales. C’est ce qu’a testé William Cooper Jr sur 66 espèces de lézards. Il a alors montré que les espèces insulaires tolèrent une plus grande proximité avec l’être humain (avant de s’enfuir) que les espèces continentales. De plus, plus l'île est loin du continent, plus le lézard tolère la proximité humaine[8]. Bien que cette expérience atteste de la docilité des espèces insulaires, elle n’en explique pas la raison.
Une telle différence pourrait s’expliquer par l’absence de prédateurs. Sans ces derniers, les espèces insulaires n’auraient pas de raison d’apprendre à fuir, en plus de risquer de perdre des ressources inutilement. En effet, la fuite est sélectionnée à condition que les individus qui ne fuient pas se fassent prédater, ce qui n’est pas le cas en absence de prédateurs. Bien que tout à fait probable, cette théorie n’est pas encore démontrée[8].
Perte de la capacité de dispersion
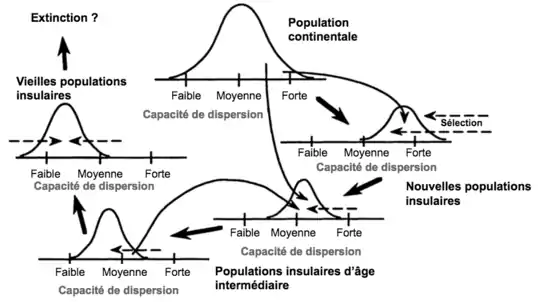
Par définition, une île est une zone propice à la vie d’une espèce entourée d’une zone qui lui est hostile, c’est pourquoi il est préférable pour ces espèces de ne pas trop se disperser au risque d'atterrir dans une zone hostile. Effectivement, il serait regrettable pour une graine d'atterrir dans l’océan car elle ne peut y pousser, c’est l’idée qu’a émise Darwin en 1855[2].
Une espèce continentale a donc une grande variabilité de dispersion possible : elle peut beaucoup se disperser ou pas beaucoup. Dans un premier temps, la dispersion d’une espèce insulaire est forte, c’est ce qui lui a permis d’atteindre l'île, mais au fur et à mesure des générations la dispersibilité diminue et devient faible[9]. Pour les raisons mentionnées précédemment, une espèce insulaire perd en capacité de dispersion au cours des générations, et au fur et à mesure que les gènes des individus qui se dispersent beaucoup, se perdent (voir Figure 2).
Exemple chez une plante anémochore
Les graines des plantes peuvent se disséminer par le vent (elles sont anémochores) ou par le biais des animaux (zoochores). Pour ce faire, elles ont développé des caractéristiques morphologiques facilitant le type de dispersion qui est présent dans l'espèce. Les plantes zoochores ont, par exemple, des crochets pour se fixer aux poils des animaux et les plantes anémochores peuvent avoir des papus qui servent de “parachutes” à la graine.
Il serait donc utile de diminuer l’efficacité de ces traits morphologiques[9], voire de les supprimer, afin de diminuer au maximum la dispersion et de s’assurer que la descendance d'une espèce se maintienne dans une zone non loin des parents, qui sont forcément situés dans une zone vivable.
Cody et Overton ont étudié l'évolution de la morphologie des propagules d’Asteraceaes (qui sont anémochores) situés sur des îlots au large de Vancouver sur plusieurs années. La taille du papus (partie permettant aux propagules de disperser) et celle de l'akène (contenant la graine) sont alors mesurées. Ainsi, si le papus diminue et que l'akène augmente, cela induit une diminution de la possibilité de dispersion, mise en avant par le ratio du volume du papule sur le volume de l'akène. D'après cette étude, la morphologie des propagules change effectivement : la taille de l'akène augmente et celle du papus diminue avec l'âge de la population. L'étude présente également une baisse significative du potentiel de dispersion avec l'augmentation de l'âge de la population (pour l’une des espèces étudiées).
Bien que des études complémentaires soient nécessaires pour affirmer qu’une évolution a bien eu lieu en faveur de la perte de dispersion, l'évolution à court terme semble la théorie la plus probables pour expliquer les modifications morphologiques observées.
Changement de fécondité
Un changement de fécondité a tout particulièrement été observé chez les oiseaux insulaires. En effet, leurs sites de nidification sont bien plus variés sur les îles que sur les continents, ce qui signifie que les lieux et les supports choisis par les oiseaux pour installer leurs nids sont plus diversifiés[1] - [2]. De plus, en milieu insulaire, la taille de la ponte diminue, cela signifie que le nombre d'œufs pondus par les oiseaux insulaires est inférieur à celui des oiseaux continentaux. Cela peut être induit par une grande diversité de raisons et différentes hypothèses s’affrontent.
- La première est une hypothèse de réallocation de l'énergie proposée par Cody[2]. Il est admis que les changements environnementaux sont plus facilement prévisibles sur les îles, ce qui rend la vie plus facile et diminue la mortalité. Ainsi, l'économie d'énergie réalisée sur la survie peut être allouée pour améliorer la qualité de la progéniture ou la longévité des parents. Il n'y a donc pas besoin de faire beaucoup de descendants pour espérer en avoir qui survivent aux modifications environnementales. Ils peuvent donc être de meilleure qualité et en plus petite quantité, d’autant plus que les parents vivent plus longtemps.
- Une seconde hypothèse proposée par Ashmole[2] se base également sur le fait que l'environnement insulaire est moins dur. Les fluctuations entre les saisons sont moins importantes que sur le continent, donc la prévisibilité des ressources est plus grande, ce qui amène à une plus petite taille de ponte en milieu insulaire.
- L'hypothèse de compensation de densité[2] se base sur le fait que la densité importante au sein d’une même espèce pourrait diminuer la taille de ponte limitant ainsi la compétition intraspécifique.
- L’absence de parasites et de prédateurs sur les îles permet une diminution du danger pour les jeunes et donc leur mortalité. Ainsi, les parents n’ont pas besoin de faire beaucoup de jeunes pour s'assurer de la viabilité de leur descendance[2].
- Enfin, la dernière hypothèse est une hypothèse de défense[2]. Étant donné les conditions de vie encore une fois plus souples et l’absence de prédateurs, le taux de survie est plus important, donc la quantité d'individus sans territoire aussi, augmentant la compétition pour accéder à un territoire. Ainsi, il est coûteux pour un individu de conserver son territoire et de le défendre face à un individu qui voudrait se l’approprier. C’est pour diminuer ce coût à la défense, que les territoires peuvent être plus petits que sur les continents et peuvent également se chevaucher. Ainsi, l'énergie économisée par cette nouvelle distribution de territoires est allouée à l'élaboration de descendants moins nombreux mais plus compétitifs.
Toutes ces hypothèses expliquant la petite taille de ponte des oiseaux insulaires sont recevables, mais à l’heure actuelle, aucune ne semble faire consensus.
Sources et références
- (en) Jacques Blondel, « Evolution and ecology of birds on islands: Trends and prospects », Vie et Milieu, vol. 50, , p. 205‑220.
- Robert J. Whittaker et José María Fernández Palacios, « Arrival and change », dans Island Biogeography: Ecology, Evolution, and Conservation, Oxford University Press, (ISBN 9780198566113, OCLC 470717243), p. 416
- (en) Mark V. Lomolino, « Body size evolution in insular vertebrates: generality of the island rule », Journal of Biogeography, vol. 32, no 10, , p. 1683‑1699 (DOI 10.1111/j.1365-2699.2005.01314.x).
- (en) Robert J. Whittaker et José María Fernández Palacios, « Speciation and the island condition », dans Island Biogeography: Ecology, Evolution, and Conservation, Oxford University Press, (ISBN 9780198566113, OCLC 470717243), p. 416.
- Christophe Terzian et Christian Biémont, « Les théories de l’effet fondateur, Mayr, et la génétique des populations », Génétique, sélection, évolution, vol. 20, no 1, , p. 111 (DOI 10.1186/1297-9686-20-1-111).
- (en) Robert P. Anderson et Charles O. Handley JR., « Dwarfism in Insular Sloths : Biogeography, Selection, and Evolutionary Rate », Evolution, vol. 56, no 5, , p. 1045‑1058 (DOI 10.1111/j.0014-3820.2002.tb01415.x).
- (en) Michael Benton et al., « Dinosaurs and the island rule: the dwarfed dinosaurs from Hatseg island », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 293, nos 3-4, , p. 438-454 (DOI 10.1016/j.palaeo.2010.01.026)
- (en) Ed Yong , « Islands make animals tamer », Nature, (DOI 10.1038/nature.2014.14462).
- (en) Martin L. Cody et Jacob McC. Overton, « Short-Term Evolution of Reduced Dispersal in Island Plant Populations », Journal of Ecology, vol. 84, no 1, , p. 53‑61 (DOI 10.2307/2261699, JSTOR 2261699).