Zoé (biologie)
Les crustacés naissent généralement sous forme de larves dont la morphologie (et éventuellement l’écologie) diffère fortement de celle des adultes. Si l'on excepte les crevettes pénéides (Dendrobranchiata) qui ont une larve nauplius et certaines espèces, d’eau douce notamment, peu nombreuses, dont le développement est direct, les crustacés décapodes européens (crabes, crevettes, pagures, homards, etc.) qui incubent leurs œufs (Pleocyemata) naissent sous forme de larves zoé ou de larves qui peuvent se rattacher à ce type.

Bref historique
En l’an X (1802), Louis-Augustin Bosc d'Antic[1] décrit et figure un crustacé planctonique qu’il nomme Zoe pelagica (du grec ζωή = vie) sans en reconnaître le caractère larvaire (cet animal avait été observé précédemment par le naturaliste hollandais Slabber en 1778[2]). En , Thompson[3] observant l’éclosion des œufs du tourteau (Platycarcinus pagurus) constate que les larves libérées ont une morphologie semblable à celle de l’organisme décrit par Bosc et en conséquence les baptise « larves zoé ». Leur forme est tellement différente de celle des adultes que cette description est accueillie avec beaucoup de scepticisme. « … Nous avons bien de la peine à croire que ces petits êtres puissent devenir des Tourteaux », dira Henri Milne Edwards[4]. Thompson[5] décrit ensuite une larve zoé étant probablement un crabe vert (Carcinus maenas) et le stade larvaire qui lui succède : la mégalope dont la locomotion est abdominale.
Sens premier de zoé
Initialement zoé désigne donc une forme larvaire de crabes. C’est un organisme planctonique dont la tête (céphalon) porte des yeux composés pairs et cinq paires d’appendices fonctionnels (antennules, antennes, mandibules, maxillules et maxilles) dont la forme est cependant bien différente de celle des appendices des adultes. Cette tête émet vers l’arrière une carapace qui recouvre le thorax et porte une épine antérieure (rostrale), une épine dorsale et, généralement, deux épines latérales. Le thorax (péréion) ne possède que deux paires d’appendices fonctionnels, le premier et le second maxillipède, qui sont biramés et dont la rame externe (exopodite) est munie de soies plumeuses, natatoires. Cette larve zoé a donc une locomotion thoracique. Les autres appendices thoraciques (du troisième maxillipède à la cinquième patte marcheuse) sont à l’état de bourgeons immobiles et les métamères correspondants très courts. L’abdomen (pléon) allongé est constitué de 5 métamères individualisés, dépourvus d’appendices fonctionnels, le sixième est encore soudé au telson équipé de fortes épines.
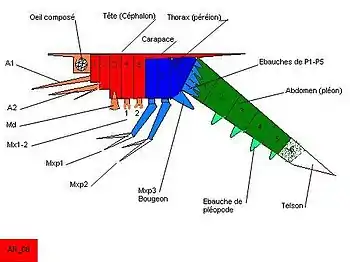
Cette larve mue plusieurs fois en se modifiant progressivement (les bourgeons des appendices thoraciques et abdominaux s’allongent notamment) mais conserve la même allure générale. Les pattes marcheuses ne portent jamais d’exopodites. On appelle stade la forme larvaire comprise entre deux mues. Ces stades sont numérotés souvent à l’aide de chiffres romains : zoé I, zoé II… L’ensemble des stades du même type constitue une phase : on parle donc de la phase zoé.
Chez les crabes notamment la première zoé (zoé I) est précédée par une prézoé. Cette larve, revêtue d’une enveloppe cuticulaire bien différente de la zoé n’est cependant qu’une forme éphémère qui mue dans les minutes (moins d’1 minute à 47 minutes[6]) qui suivent l’éclosion et passe souvent inaperçue.
Sens élargi
La grande diversité morphologique des larves de Crustacés, même chez les seuls décapodes, a conduit divers auteurs à leur donner des noms différents à en fonction de leurs particularités. Il en est résulté une certaine complexité, voire confusion, dans la terminologie. Gurney[7] propose de désigner par le nom « zoé » toutes les larves de décapodes ayant une locomotion thoracique (réalisée par les exopodites d’un nombre variable d’appendices du thorax). Elles s’intercalent entre les larves nauplius et protozoé, à locomotion céphalique, et les post-larves à locomotion abdominale. Suivant cette proposition les larves appelées antérieurement « mysis » chez les crevettes caridea sont actuellement appelées zoés. Ces larves de crevettes caridea possèdent des exopodites natatoires aux trois maxillipèdes au stade zoé I et en acquièrent ensuite sur un nombre variable de pattes thoraciques locomotrices : sur la première seulement pour la crevette grise Crangon crangon[8], sur les pattes 1 à 4 chez la crevette rose, Palaemon serratus[9] - [10].
Cependant les situations sont loin d’être toujours parfaitement tranchées et chez les crevettes pénéides les appellations de « protozoé » et de « mysis » subsistent souvent. Les protozoés ont en fait une locomotion mixte, céphalique et thoracique, par contre les mysis ont une locomotion thoracique seule. Par ailleurs on observe que chez les protozoés les yeux sont sessiles au stade I, pédonculés au stade II, et enfin que les uropodes apparaissent au stade III. Ces caractéristiques s’observent en général dans les trois premiers stades des zoés (mais celles des brachyoures n’ont pas toutes des uropodes !). Selon les auteurs les protozoés et mysis des pénéides peuvent être incluses dans la phase zoé[11] ou maintenus séparés, les mysis étant appelées zoé[12].
Larves de décapodes particulières
On continue généralement à appeler les larves de homard « mysis » parce qu’elles possèdent des exopodites natatoires sur les cinq dernières pattes thoraciques. Les maxillipèdes par contre n’ont plus de fonction locomotrice. Les trois premiers stades présentent la succession yeux sessiles au stade I, yeux pédonculés au stade II et uropodes présents au stade III. Ces larves ont donc des affinités très fortes avec les « zoés ».
Les larves des langoustes conservent également leur nom de phyllosome. Elles ont aussi des affinités zoéennes par la structure et la fonction de leurs appendices thoraciques.
Ces deux types de larves sont appelées « zoé » par Williamson[11].
D’une manière générale, les larves de Crustacés n’entrent pas aisément dans des catégories bien définies valables pour l’ensemble des espèces même dans un groupe taxonomique délimité.
Biologie
Les larves zoé sont des organismes planctoniques. Elles se nourrissent d’algues unicellulaires et d’animalcules, également planctoniques (« planctotrophie »). Les algues seules sont généralement insuffisantes pour assurer leur développement qui nécessite un apport d’origine animale. Dans les élevages artificiels les larves nauplius d’Artemia salina leur sont généralement fournies[13].
Le grand développement de l’abdomen, alors que l’organogenèse de la partie postérieure du thorax est mise en attente, peut sembler surprenant. Elle doit présenter un avantage sélectif dans le milieu planctonique.
L’abdomen, fortement musculeux, terminé par un telson élargi, particulièrement chez les crevettes Caridea, provoque, lors d’une brusque contraction, un mouvement de recul susceptible de permettre d’échapper à un prédateur. Par ailleurs, l’abdomen et le telson rabattus contre la face ventrale du céphalothorax peuvent entraîner et maintenir des proies au contact de la région buccale et même participer, par des mouvements d’extension, à leur dilacération. Enfin, l’abdomen et le telson, particulièrement chez les zoés de brachyoures, constituent un organe de toilettage qui débarrasse les appendices des éléments qui y adhèrent. La présence des épines constitue également un élément dissuasif vis-à-vis des prédateurs[14].
Les moyens de locomotion des zoés sont limités et elles paraissent prisonnières de la masse d’eau dans laquelle elles vivent. Cependant, grâce à des migrations verticales de faible amplitude (quelques mètres) elles peuvent bénéficier de courants de directions diverses et ainsi rester dans des régions aux caractéristiques compatibles avec leur développement et la vie des adultes. Un exemple classique est représenté par les zones estuariennes où les premiers stades zoé vivant en surface sont entraînés vers le large, alors que les stades plus avancés, se rapprochant du fond, sont transportés par le courant vers la partie amont.
Galerie
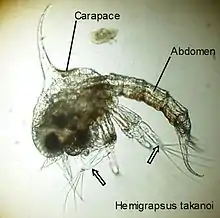 Larve zoé I du crabe Hemigrapsus takanoi. Les flèches indiquent les exopodites natatoires des maxillipèdes 1 et 2.
Larve zoé I du crabe Hemigrapsus takanoi. Les flèches indiquent les exopodites natatoires des maxillipèdes 1 et 2. Larve zoé I de Porcellana platycheles.
Larve zoé I de Porcellana platycheles.
Références
- (fr) Bosc, L. A. G. Histoire naturelle des Crustacés, contenant leur description et leurs mœurs, avec figures dessinées d’après nature. T 2, p. 135, pl. 15, Imprimerie Guilleminet Paris.
- (en) Gore, R. H. Molting and growth in decapod larvae. In : Crustacean issues, 2. Larval growth. Wenner A. M. ed. Balkema/Rotterdam/Boston.
- (en) Thompson, J. V. 1828. Zoological researches. Memoir I. On the metamorphoses of the Crustacea, and on Zoea, exposing their singular structure and demonstrating that they are not, as has been supposed, a peculiar Genus, but the larva of Crustacea. 11 p.
- Milne Edwards, 1837. Histoire naturelle des Crustacés. T2, p. 436. Imprimerie et Fonderie de Fain, Paris.
- (en) Thompson, J. V., 1835. On the double metamorphosis in the Decapodous Crustacea, exemplified in Cancer maenas, Linn. Phil. Trans London : 359-362.
- Hong S. Y. 1988. The prezoeal stage in various decapod crustaceans. Journal of Natural History, 22 : 1041-1075.
- Gurney R. 1942. Larvae of Decapod Crustacea. Ray Society, London.
- Gurney, A. 1982. The larval development of Crangon crangon (Fabr. 1795) (Crustacea : Decapoda) Bull. Br. Mus. Nat. Hist. (Zool.) 42 : 247-262.
- Fincham A. A. 1983. Larval development of British prawns and shrimps (Crustacea : Decapoda : Natantia) 4. Palaemon (Palaemon) serratus (Pennant., 1777) and functional morphology of swimming. Bull. Br. Mus. Nat. Hist. 44 : 125-161.
- Yagi, H. 1986. Le développement larvaire de la crevette rose Palaemon serratus (Pennant) (Palaemonidae, Decapoda, Crustacea) Mesogée. 46 : 29-43.
- Williamson, D. I., 1982. Larval morphology and diversity. In : The biology of Crustacea. D. E. Bliss ed. I Chief. Academic Press. Pp. 43-110.
- Schram, F. R., 1986, Crustacea. Oxford University Press. 606 p.
- Mc Conaugha, J. R. 1985. Nutrition and larval growth. In : Crustacean Issues 2. Larval growth. Wenner, A. M. ed. Balkema, Rotterdam. Pp 127-154.
- Morgan, S. G. 1987. Morphological and behavioral antipredatory adaptations in decapod zoeae. Oecologia, 73 : 393-400.