Sélection multi-niveaux
En biologie, la théorie de la sélection multi-niveaux, ou sélection multipolaire défend l’idée que la sélection naturelle est une résultante de plusieurs composantes appelées “unités de sélection”. Après la théorie de l’évolution selon le naturaliste britannique Charles Darwin, centrée sur l’individu, d’autres scientifiques ont développé différentes échelles d'interprétation de la sélection naturelle (génétique, groupe, etc.). Les interactions entre les différentes unités de sélection se traduisent par une dynamique d’où peuvent émerger des conflits.
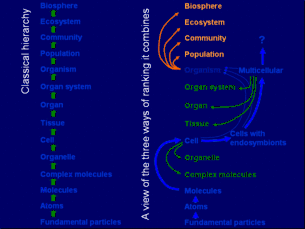
Historique
L'idée que les groupes peuvent subir des pressions de sélections comme une unité existe depuis Darwin. Les pressions de sélection ont longtemps été considérée comme axée sur l'individu. Cette vision fut de plus en plus critiquée et la vision du groupe comme entité de sélection défendue par D.S. Wilson et Sober s'est répandue et donne naissance à la notion de sélection multi-niveaux[1].
Parallèlement, Dawkins défend l'idée de "gènes égoïstes" amenant une autre vision de la sélection naturelle, plutôt gène centrée.
Certains scientifiques pensent alors qu'il faudrait voir la structure de la sélection naturelle comme une multitude d'unités de sélection. Cette conception se trouve au cœur des publications de Denis Buican depuis 1969[2]. Il la nomme sélection multipolaire et lui consacre un chapitre entier de son livre essentiel publié en 1989: la Révolution de l'évolution[3]. On pense alors que la sélection peut avoir lieu à n'importe quel niveau du vivant si trois critères sont présents : il faut une différence entre les éléments, cette différence doit donner un avantage sélectif et elle doit être transmissible à la génération suivante[4]. Plusieurs unités de sélection ont été proposées mais les trois niveaux de sélection les plus connus sont le gène, l'individu et le groupe[5].
Plus récemment, les scientifiques tentent de comprendre et de formaliser les interactions entre toutes les unités de sélection. Ainsi une unité de sélection pourrait être "désactivée" par une unité de sélection plus haute (comme l'individu qui régule la croissance de ses cellules)[5]. Wilson compare les unités de sélection aux poupées russes[6] s'emboîtant les unes dans les autres et ayant des interactions les unes avec les autres.
Sélection au niveau génétique
La sélection se fait en partie au niveau du gène. Certaines séquences ADN tels que les transposons ou encore les séquences microsatellites sont capables de se répliquer. Elles ne profitent pas à l’individu mais sont quand même répliquées au sein du génome au cours des générations, on a donc bien une sélection au niveau du gène. Richard Dawkins dans son livre Le gène égoïste [7] nous explique la façon dont le gène se réplique afin d’assurer sa survie. Le gène est « immortel » il va se répliquer et subsister à la mort de l’individu puisque ce dernier se transmettra de génération en génération. A contrario l’individu lui, n'est pas immortel. Il a une période de vie donnée.
Sélection de parentèle
La théorie de la sélection de parentèle est présentée pour la première fois par William Donald Hamilton[8] - [9] en 1964, et tend à expliquer l'ensemble des manifestations naturelles, de l'égoïsme, l'altruisme et de la malveillance. Elle représente un bon exemple de sélection au sein des gènes. En effet, on observe un comportement altruiste entre individus ayant un fort taux d'apparentement quand br>c, c'est-à-dire quand les bénéfices du receveur sont supérieurs au coût pour le donneur. L'individu cherche à transmettre ses propres allèles, que ce soit par l'intermédiaire de sa propre reproduction ou par celle des individus qui lui sont apparentés.
D'autre part, Charles Darwin, dans son ouvrage L'origine des espèces[10] fut, dans un premier temps, dubitatif face à l'organisation d'une population de fourmis. Les ouvrières ne peuvent se reproduire mais aident leur reine mère en la nourrissant et en apportant des soins aux jeunes. En effet, les ouvrières étant apparentées à la reine mère et étant plus apparentées aux autres ouvrières qu'à de potentiels descendants, il devient plus avantageux pour elles de favoriser l'aide aux autres ouvrières et à la reine par rapport à la reproduction individuelle pour assurer la transmission de leurs allèles.
Conflit gène - individu
Un conflit entre niveaux de sélection se traduit par le fait qu'un caractère étant avantageux à l'un des niveaux de sélection, ne le soit pas à un autre.
Prenons l'exemple d'un des conflits sélectifs qui concerne la mutation "t" chez la souris. La forme mutante de l'allèle "t" est portée par les cellules sexuelles de la souris. Cette forme mutante diminue la viabilité des autres gamètes, et assure ainsi sa transmission dans les générations suivantes même si elle n'est présente que dans la moitié des cellules sexuelles. Le conflit ici est traduit par le fait que cet allèle, si présent en double exemplaire chez un individu homozygote, présente un effet phénotypique létal, entraînant presque systématiquement l'avortement de l'embryon. Cet exemple montre comment un gène extrêmement désavantageux pour l'espèce se trouve pourtant favorisé par la sélection, grâce au fait qu'il augmente le nombre de ses copies à la génération suivante. La valeur sélective de cette mutation peut être considérée de deux points de vue différents : si on la considère au niveau du gène, cette mutation "t" est avantageuse, puisque le nombre de ses copies augmente à chaque génération, mais si on la considère au niveau de l'individu, c'est une mutation désavantageuse puisqu'elle conduit à l'avortement de l'embryon, et donc à un nombre de descendants moindre[11].
Conflit intra - génomique
Prenons l'exemple des transposons: ceux-ci, bien que n'ayant pas de rôle bien précis, se répliquent et s'insèrent sur la séquence ADN de certains gènes. Leurs réplications peuvent entraîner des mutations ayant un effet délétère sur l'individu, et donc agir sur sa fitness. Mais ils peuvent aussi empêcher l'expression des gènes sur lesquelles ils se sont répliqués, leur causant préjudice.
Conflit inter - génomique
On constate parfois des conflits entre gènes de génomes différents. Certains parasites comme sacculina carcini, féminisent leurs hôtes. Elles parasitent majoritairement les crabes mâles dont l'abdomen se féminise. Ce procédé permettrait aux parasites d’augmenter leur longévité. Il y a donc bien une modification de l’expression des gènes du crabe.
Sélection au niveau individuel
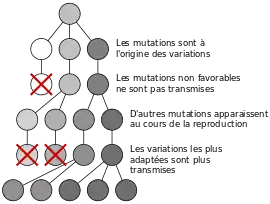
C'est le premier niveau de sélection établi à la suite de l'élaboration de la théorie de sélection naturelle. Selon Darwin la sélection s'effectue au niveau de l'individu selon le principe de la sélection naturelle : un individu dont les traits de caractère lui confèrent un avantage sélectif en termes d'accès aux ressources, de survie ou de fécondité (fitness ou valeur sélective) sera avantagé par rapport aux autres individus de la même espèce. Il l'explique ainsi dans l'origine des espèces :
" En nous rappelant le fait qu'il nait toujours beaucoup plus d'individus qu'il ne peut en vivre, pouvons nous douter que les individus possédant un avantage quelconque[...] aient la meilleure chance de se reproduire? [...] toute variation, si peu nuisible qu'elle soit à l'individu, entraîne forcément la disparition de celui-ci. J'ai donné le nom de sélection naturelle, ou de persistance du plus apte à cette conservation des différences et des variations individuelles favorables et à cette élimination des variations nuisibles."[10]
Cela favorise alors l'individu avec la plus forte fitness, c'est-à-dire la capacité d'un individu à se reproduire.
Groupe ou individu ?
La question du niveau auquel agit le processus de sélection naturelle est, depuis les années 1960, sujette à de nombreux débats. Le biologiste Vero Copner Wynne-Edwards propose, en 1962, une hypothèse selon laquelle la plupart des espèces animales maintiennent la densité de leur population à un niveau optimal compte tenu de leur habitat. Dont l'explication serait que l'individu régule par lui-même son impact sur les ressources du milieu afin de favoriser la survie du groupe. Vision notamment attaquée par George C. Williams qui propose une vision plus individualiste du mécanisme. Le comportement optimise l'individu et la sélection n'opère qu'à ce seul niveau. Controverse stoppée par un consensus : la sélection de groupe est une possibilité théorique, les conditions de mise en pratique sur le plan empirique sont extrêmement contraignantes. On estime alors que la sélection naturelle ne peut agir qu'à un seul niveau l'individu. C'est en 1976 seulement que cela est remis en question de nouveau par Richard Dawkins avec la thèse « le gène égoïste » qui révèle un nouveau niveau de sélection : le gène. Plaçant l'individu comme un simple véhicule[12].
Sélection au niveau du groupe
Historique

En 1871 Charles Darwin, dans son ouvrage « The Descent of Man », évoque l'existence d'un comportement solidaire entre individus d'un même groupe, et le compare au processus de civilisation :
"It must not be forgotten that although a high standard of morality gives but a slight or no advantage to each individual man and his children over the other men of the same tribe, yet that an increase in the number of well-endowed men and advancement in the standard morality will certainly give an immense advantage to one tribe over another. There can be no doubt that a tribe including many members who, from possessing in a high degree the spirit of patriotism, fidelity, obedience, courage, and sympathy, were always ready to aid one another, and to sacrifice themselves for the common good, would be victorious over most other tribes ; and this would be natural selection."[13]
Il s'agirait bien d'une sélection naturelle basée sur l’intérêt qu’un individu porte pour la fitness du groupe, au détriment de sa propre fitness.
L’évocation d’une sélection favorisant la fitness du groupe mène à un grand débat, puisque souvent, seule la sélection aux niveaux génétique et individuel étaient considérées dans le cadre de la sélection naturelle[13].
G.C Williams, en 1966, dans son ouvrage Adaptation and Natural Selection, critique fortement l'existence d'une solidarité des individus au sein d'un groupe et l'hypothèse selon laquelle il existerait peut-être un mécanisme de sélection de groupe dépassant le cadre de la parentèle, et qui fonctionnerait sur le modèle de la sélection naturelle[14].
Et c'est 1975, la théorie de la sélection au niveau d'un groupe dans son ensemble plutôt qu'au niveau de chacun des individus composant le groupe est proposée de nouveau, cette fois par David S. Wilson. L'idée de base de cette théorie est qu'un trait pouvant être défavorable à un individu peut néanmoins être transmis dans une population si ce trait confère un avantage au groupe dont fait partie l'individu, par rapport à d'autres groupes.
Définition
La sélection de groupe est donc un mécanisme évolutif dans laquelle la sélection naturelle a un impact au niveau du groupe et non pas au niveau de l'individu. Un tel groupe possède donc des individus ayant chacun un certain statut social, entraînant une certaine structure qui permet au groupe d'avoir une meilleure fitness.
Altruisme
Un comportement altruiste traduit la volonté d’un individu au sein d’un groupe à reduire sa propre fitness (par des comportements handicpants du point de vue individuel) pour le bien de celle d’un ou plusieurs autres individus du groupe. Ce trait sera sélectionné aux générations suivantes car il permet au groupe d'avoir une meilleure fitness.
En 1964, Hamilton défini la fitness inclusive et la présente comme la raison de la coopération dans les colonies d'insectes. Selon la règle d'Hamilton, on observera un comportement altruiste si :
où correspond au bénéfice du receveur, à ses couts, et le degré d'apparentement entre le donneur et le receveur. Cependant des recherches ont montré que cette règle était incomplète (voire fausse comme dans le cas où r=0)[15].




D’après E. O. Wilson, si l'on considère seulement cette équation, certaines colonies n'ont pas le comportement maximisant leurs fitness car l’équation ne prend pas en compte la sélection au niveau de la colonie (sélection au niveau de la metapopulation). En effet, une population altruiste a plus de chance d'augmenter en fréquence qu'une population non altruiste (voir metapopulations). La formule proposé est avec qui correspond au bénéfice du receveur et correspondant au bénéfice lié à la sélection au niveau de la metapopulation[15].



On peut parler ici de conflit groupe-individu car un trait désavantageux au niveau de l'individu (l'altruisme, qui réduit la fitness de l'individu) est pourtant sélectionné puisqu'il rapporte une meilleure fitness au niveau du groupe. Il n'y a cependant pas toujours conflit et un trait favorable au groupe peut aussi être favorable à l'individu[16].
Altruisme réciproque
.jpg.webp)
Une autre forme d'altruisme est également constatée chez certaines espèces animales. Il s'agit de l'altruisme réciproque, théorie proposée par le biologiste américain Robert Trivers en 1971, qui serait une forme de coopération mutuelle.
Les populations de chauves-souris vampires constituent un exemple d'altruisme réciproque. En effet, ces chauves-souris qui vivent en groupe de 8 à 12 individus, meurent en cas de très faibles ressources alimentaires. Pour éviter la mort d'individus de leur groupe par dénutrition, les chauves-souris qui trouvent une proie aspirent une grande quantité de sang dont ils partagent une partie avec un autre membre de leur groupe. Ce partage se fait de manière réciproque avec les autres individus, qu'ils soient apparentés ou non. Mais si l'un des individus ne veut pas partager sa nourriture avec d'autres membres alors qu'il a déjà reçu de la nourriture de la part des autres, (il est donc considéré comme "tricheur" car il refuse la réciprocité), il sera exclu du prochain partage de nourriture[17].

Être exclu d'un partage augmente les risques de la dénutrition et donc de la mort de l'individu. Ainsi, partager son alimentation est un moyen d'assurer sa survie en même temps que la survie des autres, puisque l'individu qui partage réduit donc sa propre alimentation pour permettre aux autres de vivre. Ce caractère altruiste sera également favorisé par la sélection car il amène une meilleure fitness du groupe.
Les suricates sont également un exemple de population animale faisant preuve d'altruisme réciproque. En effet, une population de suricates est composée de quelques sentinelles qui montent la garde pour le groupe entier, et préviennent les autres des dangers extérieurs ou de la présences d'éventuels prédateurs, mettant en danger leurs propres vies. Mais chaque individu du groupe peut à son tour devenir une sentinelle et protéger les autres. Les individus d'une population de suricate n'ont pas de statut social fixé.
Autres niveaux de sélection
D'autres unités de sélection moins connues existent.
Niveau cellulaire
Les débuts de la microscopie poussa les scientifiques a rechercher l'origine de la lutte pour la vie et de la sélection jusque dans les cellules et les molécules. Ainsi Wilhelm Roux (reprenant Lamarck) fonda sa théorie de la sélection entre cellules par la lutte des parties entre elles.
Certains théoriciens de l'hérédité avancèrent même l'idée que la sélection s'effectuait au niveau de "particules élémentaires invisibles"[18] accumulées soit dans les cellules somatiques soit dans les cellules germinales. August Weismann a conçu dans ce sens la théorie la plus complexe qu'il nomma la sélection germinale.
Une autre théorie de la sélection au niveau de la cellule a été avancé par Leo Buss dans son livre "The Evolution of Individuality". Le cancer serait alors un symptôme d'un conflit entre le niveau cellulaire et le niveau de l'individu[5] et la chimiothérapie serait une méthode pour réduire la fitness des cellules cancéreuses[19].
Dèmes ou métapopulation
La métapopulation est un ensemble fermé qui n'échange pas de gènes avec l'extérieur. La sous-population ou dème est une subdivision géographique représentée par exemple par un bois, une mare, un lac, une zone littorale. La dérive génétique va agir différemment dans chaque dème
Dans une métapopulation ou un dème, qui est un ensemble de plusieurs populations d'une même espèce séparées spatialement ou temporellement, un trait peut être sélectionné même s'il est handicapant du point de vue d'une autre unité de sélection. Pour illustrer le niveau de sélection de la métapopulation, Wilson prend l'exemple de l'altruisme[16].
Soit par exemple deux individus a et b dans un même groupe, on peut parler d’altruisme si le bénéfice pour l’individu b tiré du sacrifice de l’individu a est supérieur au coût du sacrifice de l’individu a. Un groupe possédant des individus altruistes a moins de chances de disparaître qu'un autre groupe dans lequel chaque individu ne privilégierait que son propre intérêt individuel. Les comportements altruistes seront favorisés par la sélection dans les groupes hétérogènes puisqu'ils mènent à une meilleure fitness de groupe[7] - [20] - [21].
Notes et références
- Wilson, D. S., Van Vugt, M., & O'Gorman, R. (2008). Multilevel selection theory and major evolutionary transitions implications for psychological science. Current Directions in Psychological Science, 17(1), 6-9.
- Grimoult, C. (2009), La Preuve par neuf. Les Révolutions de la pensée évolutionniste, Ellipses, p.133-149.
- Buican, D.(1989). La Révolution de l'évolution, Paris, PUF.
- Wade, M. J. (1982). Evolution of interference competition by individual, family, and group selection. Proceedings of the National Academy of Sciences, 79(11), 3575-3578.
- Gould, S. J., & Lloyd, E. A. (1999). Individuality and adaptation across levels of selection: How shall we name and generalize the unit of Darwinism?. Proceedings of the National Academy of Sciences, 96(21), 11904-11909.
- Wilson, D. S.; Wilson, E. O. (2008). "Evolution 'for the good of the group'.". American Scientist 96 (5): 380–389. doi:10.1511/2008.74.1.
- Richard Dawkins, Le gène égoïste, Angleterre, Odile Jacobs, , 459 p. (ISBN 978-2-7381-1243-9)
- Hamilton, W.D, « The Genetical Evolution of Social Behaviour. I », Journal of Theoretical Biology, no 7, , p. 1-16
- Hamilton, W.D, « The Genetical Evolution of Social Behaviour. II », Journal of Theoretical Biology, no 7, , p. 17-52
- Charles Darwin, On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, Londres, John Murray,
- Buican Denis et Grimoult Cédric, L'évolution : Histoire et Controverses, Paris, CNRS Editions, , 246 p. (ISBN 978-2-271-07223-8)
- Cyril Hédoin (2012). Veblen et son actualité : sélection multi-niveaux et darwinisme généralisé. OEconomia, 2012, p. 179-207 doi:10.4074/S2113520712012030
- (en) Charles Darwin, The Descent of Man, and Selection in Relation to Sex, United Kingdom, John Murray,
- (en) George C. Williams, Adaptation and Natural Selection : a critique of some current evolutionary thought, Princeton, N.J., Princeton University Press, , 307 p. (ISBN 0-691-02615-7)
- Wilson, E. O. (2005). "Kin Selection as the Key to Altruism: its Rise and Fall". Social Research 72 (1): 159–166
- Wilson, D.S. (1975). A theory of group selection. Proc Natl Acad Sci U S A, 72, 143-146
- Peter H. Raven, Georges B. Johnson, Kenneth A. Mason, Jonathan B. Losos, Susan S. Singer (trad. de l'anglais), Biologie, Bruxelles/Paris, De Boeck, , 1406 p. (ISBN 978-2-8041-6305-1)
- H Lebrun (1912) : Néo-Darwinisme et Néo-Lamarckisme, Revue néo-scolastique de philosophie, numéro 76, p. 489-514
- Lewontin, R. C. (1970). The units of selection. Annual Review of Ecology and Systematics, 1-18.
- (en) Jeffrey A. Fletcher, Martin Zwick, « The Evolution of Altruism : Game Theory in Multilevel Selection and Inclusive Fitness », Journal of Theoretical Biology, no 245, , p. 26-36
- (en) Matthijs Van Veelen, « Group Selection, Kin Selection, Altruism and Cooperation: when inclusive fitness is right and when it can be wrong », Journal of Theoretical Biology, no 259, , p. 589-600
Annexes
Articles connexes
Bibliographie
- Cyril Hédoin (2012). Veblen et son actualité : sélection multi-niveaux et darwinisme généralisé. OEconomia, 2012, p. 179-207 doi:10.4074/S2113520712012030
- Lewontin, R. C. (1970). The units of selection. Annual Review of Ecology and Systematics, 1-18.
- Buican D., Grimoult C. (2011). L'évolution : Histoire et controverses, CNRS Éditions
- Mérat. P (1969) p. 243-272 ann. Génét. Sél. anim.
- Brondex. F (1999) Évolution, synthèse des faits et théories, édition DUNOD (ISBN 2-10-004006-5)