Phototropisme
Le phototropisme est la capacité d'un organisme (ex: plantes, champignons) à s'orienter par rapport à la lumière[1]. Ce tropisme, distinct du phototactisme, s'effectue dans la direction de la lumière.



Les scientifiques distinguent plusieurs phototropismes en fonction de l'orientation de la lumière[2] :
- Phototropisme positif ou organisme phototrope positif : croissance dans la direction de la lumière. Exemple : tiges.
- Phototropisme négatif ou organisme phototrope négatif : dans la direction opposée. Exemple : racines.
- Orthophototropisme ou organisme orthophototrope ou orthophototropique : dans la direction parallèle. Exemple : tiges et coléoptiles.
- Diaphototropisme ou organisme diaphototrope ou diaphototropique : dans la direction perpendiculaire . Exemple : feuilles.
Lorsque le stimulus est un rayonnement solaire, les scientifiques parlent plus spécifiquement d’héliotropisme, mais ce terme est devenu désuet[2].
En biologie, certaines espèces ou certains organes font preuve d'un phototropisme marqué. C'est le cas des organes aériens des végétaux.
Historique de la découverte et compréhension du phototropisme
Une expérience historique pour démontrer l'existence du phototropisme consiste à laisser germer une pomme de terre à l'intérieur d'une sorte de labyrinthe, avec une seule source de lumière. Au cours de la croissance, la plante suivra le plus court chemin vers la source de lumière. L'intérêt fonctionnel du phototropisme est de permettre aux plantes d'accéder au meilleur ensoleillement possible permettant ainsi d'assurer la photosynthèse de façon optimale.
Le phototropisme a été pour la première fois décrit par Darwin qui avait démontré ce phénomène avec des coléoptiles de poacées.
Il a ensuite été montré chez les plantes que certains organes et une hormone spécifique, l'auxine jouent un rôle important dans le phototropisme.
Rôle de l'auxine
Les auxines induisent l'élongation des cellules des plantes supérieures et sont produites dans la partie apicale des végétaux. Elles suivent généralement un transport basipète dans les tissus. Le modèle de Cholodny–Went explique qu'en présence d'un éclairage anisotrope, ce transport d'auxine vertical est modifié. Un transfert horizontal d'auxine a lieu dans la direction de la partie la moins éclairée. Ceci induit une plus forte concentration d'auxine dans la partie non éclairée, qui s'allonge donc davantage que la partie éclairée. Il en résulte une incurvation de la plante vers la source de lumière (phototropisme positif).
La plupart des pousses de plantes présentent un phototropisme positif et réorganisent leurs chloroplastes dans les feuilles pour maximiser l'énergie photosynthétique et favoriser la croissance[3] - [4]. Certaines pointes de pousses de vigne présentent un phototropisme négatif, ce qui leur permet de se développer vers des objets sombres et solides et de les escalader. La combinaison du phototropisme et du gravitropisme (la capacité d'un organisme à s'orienter par rapport à la gravité) permet aux plantes de pousser dans la bonne direction[5].
Mécanisme
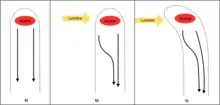
Il existe plusieurs molécules de signalisation qui aident la plante à déterminer d'où vient la source de lumière. Celles-ci activent plusieurs gènes, qui modifient les gradients hormonaux permettant à la plante de se développer vers la lumière. La pointe même de la plante est connue sous le nom de coléoptile, qui est nécessaire à la détection de la lumière[3]. La partie médiane du coléoptile est la zone où se produit la courbure des pousses. L'hypothèse Cholodny-Went, développée au début du 20e siècle, prédit qu'en présence de lumière asymétrique, l'auxine se déplacera vers le côté ombragé et favorisera l'allongement des cellules de ce côté pour faire courber la plante vers la source lumineuse[6]. Les auxines activent les pompes à protons, diminuant le pH dans les cellules du côté obscur de la plante. Cette acidification de la région de la paroi cellulaire active des enzymes appelées expansines qui perturbent les liaisons hydrogène dans la structure de la paroi cellulaire, rendant les parois cellulaires moins rigides. De plus, une activité accrue de la pompe à protons conduit à plus de solutés pénétrant dans les cellules végétales du côté obscur de la plante, ce qui augmente le gradient osmotique entre le symplaste et l'apoplaste de ces cellules[7]. L'eau pénètre alors dans les cellules le long de son gradient osmotique, entraînant une augmentation de la pression de turgescence. La diminution de la résistance de la paroi cellulaire et l'augmentation de la pression de turgescence au-dessus d'un seuil de rendement[8] provoquent le gonflement des cellules, exerçant la pression mécanique qui entraîne le mouvement phototropique.
Les protéines codées par un deuxième groupe de gènes, les gènes PIN, jouent un rôle majeur dans le phototropisme. Ce sont des transporteurs d'auxine et on pense qu'ils sont responsables de la polarisation de l'emplacement de l'auxine. Plus précisément, PIN3 a été identifié comme le principal transporteur d'auxine[9]. Il est possible que les phototropines reçoivent de la lumière et inhibent l'activité de la PINOID kinase (PID), qui favorise alors l'activité de PIN3. Cette activation de PIN3 conduit à une distribution asymétrique de l'auxine, qui conduit ensuite à un allongement asymétrique des cellules de la tige. Les mutants pin3 avaient des hypocotyles et des racines plus courts que le type sauvage, et le même phénotype a été observé chez les plantes cultivées avec des inhibiteurs d'efflux d'auxine[10]. En utilisant un marquage immunogold anti-PIN3, le mouvement de la protéine PIN3 a été observé. PIN3 est normalement localisé à la surface de l'hypocotyle et de la tige, mais est également internalisé en présence de brefeldine A (BFA), un inhibiteur de l'exocytose. Ce mécanisme permet à PIN3 d'être repositionné en réponse à un stimulus environnemental. On pense que les protéines PIN3 et PIN7 jouent un rôle dans le phototropisme induit par les impulsions. Les réponses de courbure chez le mutant "pin3" ont été réduites de manière significative, mais seulement légèrement réduites chez les mutants "pin7". Il existe une certaine redondance entre "PIN1", "PIN3" et "PIN7", mais on pense que PIN3 joue un rôle plus important dans le phototropisme induit par les impulsions[11].
Il existe des phototropines qui sont fortement exprimées dans la région supérieure des coléoptiles. Il existe deux phototropismes principaux, ils sont phot1 et phot2. Les mutants simples phot2 ont des réponses phototropes comme celles du type sauvage, mais les doubles mutants phot1 phot2 ne montrent aucune réponse phototrope[5]. Les quantités de PHOT1 et PHOT2 présentes sont différentes selon l'âge de la plante et l'intensité de la lumière. Il y a une grande quantité de PHOT2 présente dans les feuilles matures d'Arabidopsis et cela a également été observé dans les orthologues du riz. L'expression de PHOT1 et PHOT2 change en fonction de la présence de lumière bleue ou rouge. Il y avait une régulation à la baisse de l'ARNm de PHOT1 en présence de lumière, mais une régulation à la hausse du transcrit de PHOT2. Les niveaux d'ARNm et de protéines présents dans la plante dépendaient de l'âge de la plante[12]. Cela suggère que les niveaux d'expression de la phototropine changent avec la maturation des feuilles. Les feuilles matures contiennent des chloroplastes qui sont essentiels à la photosynthèse. Le réarrangement des chloroplastes se produit dans différents environnements lumineux pour maximiser la photosynthèse. Il existe plusieurs gènes impliqués dans le phototropisme des plantes dont les gènes NPH1 et NPL1. Ils sont tous deux impliqués dans le réarrangement des chloroplastes[4]. Les doubles mutants nph1 et npl1 se sont avérés avoir des réponses phototropes réduites. En fait, les deux gènes sont tous les deux redondants pour déterminer la courbure de la tige.
Effets de la longueur d'onde
Le phototropisme chez des plantes comme Arabidopsis thaliana est dirigé par des récepteurs de lumière bleue appelés phototropines[13]. D'autres récepteurs photosensibles chez les plantes comprennent les phytochromes qui détectent la lumière rouge[14] et les cryptochromes qui détectent la lumière bleue. Différents organes de la plante peuvent présenter différentes réactions phototropes à différentes longueurs d'onde de lumière. Les extrémités des tiges présentent des réactions phototropes positives à la lumière bleue, tandis que les extrémités des racines présentent des réactions phototropes négatives à la lumière bleue. Les extrémités des racines et la plupart des extrémités des tiges présentent un phototropisme positif à la lumière rouge. Les cryptochromes sont des photorécepteurs qui absorbent la lumière bleue/UV-A, et ils aident à contrôler le rythme circadien des plantes et le moment de la floraison. Les phytochromes sont des photorécepteurs qui détectent la lumière rouge/rouge lointain, mais ils absorbent également la lumière bleue ; ils peuvent contrôler la floraison des plantes adultes et la germination des graines, entre autres. La combinaison des réponses des phytochromes et des cryptochromes permet à la plante de répondre à différents types de lumière[15]. Ensemble, les phytochromes et les cryptochromes inhibent le gravitropisme dans les hypocotyles et contribuent au phototropisme.
Notes et références
- Mémo visuel de biologie végétale, Malakoff, Dunod, , 201 p. (ISBN 978-2-10-084339-8), p. 89
- Romaric Forêt, Dictionnaire des sciences de la vie, De Boeck Superieur, , p. 1058
- Goyal, A., Szarzynska, B., Fankhauser C. (2012). Phototropism: at the crossroads of light-signaling pathways. Cell 1-9.
- T. Sakai, T. Kagawa, M. Kasahara, T.E. Swartz, J.M. Christie, W.R. Briggs, M. Wada et K. Okada, « Arabidopsis nph1 and npl1: Blue light receptors that mediate both phototropism and chloroplast relocation », PNAS, vol. 98, no 12, , p. 6969–6974 (PMID 11371609, PMCID 34462, DOI 10.1073/pnas.101137598
 , Bibcode 2001PNAS...98.6969S)
, Bibcode 2001PNAS...98.6969S) - Liscum, E. (2002). Phototropism: Mechanisms and Outcomes. Arabidopsis Book 1-21.
- J.M. Christie et A.S. Murphy, « Shoot phototropism in higher plants: New light through old concepts », American Journal of Botany, vol. 100, no 1, , p. 35–46 (PMID 23048016, DOI 10.3732/ajb.1200340)
- Achim Hager, « Role of the plasma membrane H+-ATPase in auxin-induced elongation growth: historical and new aspects », Journal of Plant Research, vol. 116, no 6, , p. 483–505 (ISSN 1618-0860, PMID 12937999, DOI 10.1007/s10265-003-0110-x, S2CID 23781965)
- Daniel J. Cosgrove, Elizabeth Van Volkenburgh et Robert E. Cleland, « Stress relaxation of cell walls and the yield threshold for growth: Demonstration and measurement by micro-pressure probe and psychrometer techniques », Planta, vol. 162, no 1, , p. 46–54 (ISSN 0032-0935, PMID 11540811, DOI 10.1007/BF00397420, S2CID 6870501)
- Z. Ding, C.S. Galván-Ampudia, E. Demarsy, L. Langowski, J. Kleine-Vehn, Y. Fan, M.T. Morita, M. Tasaka, C. Fankhauser, R. Offringa et J. Friml, « Light-mediated polarization of the PIN3 auxin transporter for the phototropic response in Arabidopsis », Nature Cell Biology, vol. 13, no 4, , p. 447–453 (PMID 21394084, DOI 10.1038/ncb2208, S2CID 25049558)
- J. Friml, J. Wisniewska, E. Benkova, K. Mendgen et K. Palme, « Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis », Nature, vol. 415, no 6873, , p. 806–809 (PMID 11845211, DOI 10.1038/415806a, Bibcode 2002Natur.415..806F, S2CID 4348635, lire en ligne)
- K. Haga et T. Sakai, « PIN Auxin Efflux Carriers Are Necessary for Pulse-Induced But Not Continuous Light-Induced Phototropism in Arabidopsis », Plant Physiology, vol. 160, no 2, , p. 763–776 (PMID 22843667, PMCID 3461554, DOI 10.1104/pp.112.202432 )
- J. Labuz, O. Sztatelman, A. K. Banas et H. Gabrys, « The expression of phototropins in Arabidopsis leaves: developmental and light regulation », Journal of Experimental Botany, vol. 63, no 4, , p. 1763–1771 (PMID 22371325, DOI 10.1093/jxb/ers061 )
- « Phototropins: Photoreceptors that provide a novel photochemical mechanism for signaling » [archive du ] (consulté le )
- « Phytochrome », sur plantphys.info (consulté le )
- McCoshum, S., Kiss, J.Z. (2011). Green light affects blue-light based phototropism in hypocotyls of Arabidopsis thaliana. Journal of the Torrey Botanical Society 138(4), 409-417. DOI 10.3159/TORREY-D-11-00040.1. JSTOR:41475107.
Voir aussi
Articles connexes
Liens externes
Bibliographie
- Viaud, G. (1948). Le phototropisme animal: aspects nouveaux de la question (Vol. 10). J. Vrin.