Melon (anatomie)
Le melon est un organe ovale de tissu gras occupant la majeure partie de la tête chez les Delphinidae. Chez les cachalots ou les baleines à bec, où il est très développé, il est appelé spermaceti.
Le melon du béluga est aussi unique car le béluga peut en changer la forme à volonté. La fonction du melon n'est pas totalement connue, il semble qu'il permette de détecter plus efficacement les sons utilisés dans l'écholocation, servant ainsi de sonar (en particulier pour les dauphins). Il pourrait aussi servir dans les plongées profondes comme organe régulateur de flottabilité ; il y a un lien entre l'aptitude aux plongées profondes et la taille de l'organe.
Évolution et développement
Ontogenèse

L’évolution des tissus spécialisés contribuant à la production de sons pour l’écholocation chez les Odontoceti reste encore méconnue. Une étude portant sur l’ontogenèse des structures impliquées dans la production et la modulation de sons chez les Delphinidae a été réalisée par G. Frainer et al.[1] afin de comparer les structures nasales fœtales, périnatales, néonatales et matures de spécimens de Pontoporiidae, de Phocoenidae et de Delphinidae. En identifiant les transformations morphologiques ontogéniques des tissus nasaux, il est possible de répondre à la question suivante: quels sont les processus hétérochroniques qui interviennent dans les adaptations de la production du son chez les odontocètes? Ainsi, il est possible d’avoir une meilleure compréhension phylogénétique et évolutive de ces cétacés.
L’évolution des cétacés dans l’eau a entraîné un réarrangement du crâne, du maxillaire et du prémaxillaire[2]. Ces changements ont impliqué un réarrangement des muscles et des tissus frontaux afin d’ouvrir et de fermer le passage nasal. Plusieurs de ces changements ont lieu pendant la morphogenèse du crâne et sont contrôlés par la transformation du cartilage nasal au stade fœtal[3]. Chez les odontocètes, le développement fœtal du cartilage nasal affecte le tissu nasal trouvé chez les adultes. Ainsi, en étudiant les tissus nasaux, il est possible de retracer le développement morphologique des structures impliquées dans la production de son chez différents groupes d'odontocètes.
À l’aide d’analyses microscopiques, d’image à résonance magnétique, de tomodensitométrie et de dissections, les chercheurs ont réussi à tracer l’évolution et l’origine des différentes structures produisant des sons ainsi que leurs tissus adjacents. Ils ont ainsi pu créer un arbre phylogénétique pour retracer l’évolution des structures produisant du son pour différents clades d’odontocètes.
À la naissance, toutes les espèces étudiées possèdent les structures impliquées dans la production de sons, toutefois des changements allométriques et des transformations au niveau de la composition des tissus ont lieu pour former un stade mature.
Le melon postérieur est composé de 2 masses. Afin de rendre compte de cette découverte, les chercheurs proposent de les nommer Corpus adiposum nasalis terminalis pars posterioris dexter (Cand) et Corpus adiposum nasalis terminalis pars posterioris sinister (Cans). La morphologie des deux masses du melon varie considérablement chez les Delphinoidea. De plus, le melon postérieur et le melon sont des structures distinctes issues de tissus indépendants et de facteurs de régulation différents.
Finalement, selon la reconstruction de l’arbre phylogénétique des odontocètes, l’ancêtre commun des dauphins qui se trouvaient dans la famille des Kentriodontidae possédait une morphologique unique des tissus nasaux qui serait adaptée pour des sons très dirigés et directs. L’analyse des tissus nasaux des odontocètes modernes laisse suggérer que ces derniers n’auraient pas évolué sous des contraintes concernant la direction de leurs sons et que ces sons seraient moins directs et dirigés que leurs ancêtres. La modulation des sons serait un changement évolutif important qui aurait découlé de l’évolution des structures produisant les sons chez les odontocètes modernes.
En somme, les mécanismes ontogéniques sont parmi les facteurs les plus importants pour expliquer l’évolution de la tête chez les odontocètes. La reconstruction phylogénétique permet d’affirmer que les odontocètes possèdent des systèmes spécialisés de production et d’émission de sons et que ces systèmes ont évolué quatre fois de manière indépendante sous des processus ontogéniques sur les structures de communication et d’écholocation.
Morphologie et fonctions
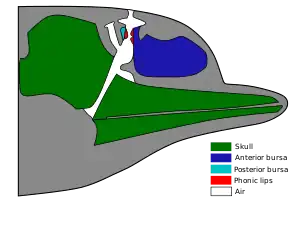
L’écholocation est un processus complexe qui implique plusieurs structures anatomiques et tissus de la tête des Odontoceti. Ces structures et tissus peuvent varier, conduisant ainsi à des différences acoustiques entre les espèces. À la lumière de ces variations interspécifiques, quelles sont les différences qualitatives et quantitative de l'anatomie du melon chez les odontocètes? Une étude menée par McKenna et al[4]. répond à cette question et permet d'extrapoler les processus d’écholocation aux espèces qui sont peu étudiées ou méconnues. Les relations géométriques du melon par rapport aux structures et tissus connexes ont été conservées afin de comprendre les fonctions bioacoustiques du melon et l’implication des tissus adjacents dans le processus d’écholocation.
La production de son chez les odontocètes commence par l’action du muscle palatopharyngien qui pousse le larynx dorsalement sur les conduits osseux des narines ce qui augmente la pression de l’air dans les narines. Cet air pressurisé passe à travers les lèvres phoniques ce qui entraîne des vibrations dans les corps adipeux de la bourse dorsale. Ces vibrations interagissent avec le crâne, le tissu conjonctif et un système de sacs d’airs nasaux qui agissent conjointement comme un miroir acoustique. Finalement, ces vibrations prennent plusieurs chemins à travers le melon et émergent dans l’environnement [5].
Lors de cette étude de McKenna et al., les melons de neuf espèces d’odontocètes regroupant cinq familles ont été étudiées à l’aide de scans tomodensitométriques à rayon X ce qui permet de conserver l’intégrité géométrique de leur anatomie. Les auteurs ont identifié les éléments anatomiques qui influencent la capacité du melon à concentrer les sons. Tout d’abord, le profil de tissus adipeux et leur changement de densité sont un indicateur de la réfraction du son à l’intérieur du melon. Ainsi, la composition et la topographie des lipides peuvent être utiles pour une espèce qui vit en profondeur, car elle ne peut se servir de ses sacs d’air pour réfléchir le son étant donné que le volume d’air est limité par la forte pression en profondeur. Ces lipides seraient donc un substitut aux espaces d’airs chez les espèces qui plongent en grande profondeur. Par la suite, la longueur du melon peut augmenter la capacité à concentrer le son. En effet, plus le trajet du son à travers les tissus adipeux est long, plus le son pourra être concentré. Les cachalots illustrent bien cette capacité, car ces odontocètes possèdent l’organe du spermaceti qui combiné avec leur melon augmentent significativement le trajet du son à travers les lipides [6]. Par la suite, la forme du melon qui varie entre les espèces pourrait aussi augmenter cette capacité à concentrer les sons. Finalement, la position des lèvres phoniques et de la bourse dorsale par rapport au melon est importante pour comprendre comment le son voyage à l’intérieur de la tête, car cette source du son varie selon les espèces.
Une autre fonction bioacoustique du melon serait d’agir comme filtre face aux sons de basse fréquence. En effet, le diamètre du melon pourrait empêcher les fréquences plus longues que la longueur du melon d’être amplifiées par celui-ci. Un des avantages reliés à cette fonction serait que les sons de basses fréquences qui voyagent plus loin auraient moins de chance d’être entendus par les prédateurs.
Il est aussi important de savoir que la taille du melon augmente avec la taille de l’animal, mais cette relation n’est pas isométrique.
En somme, les différences du melon au niveau de la taille, de la forme et de sa position sont des indicateurs du comportement du son à travers les différentes structures de la tête chez les odontocètes. Ces pistes de comparaison entre les structures anatomiques impliquées dans l’écholocation chez différents groupes d’odontocètes permettent de construire des modèles plus robustes pour comprendre les mécanismes généraux derrière l’écholocation.
Références
- Frainer, G., Moreno, I. B., Serpa, N., Galatius, A., Wiedermann, D., & Huggenberger, S. (2019). Ontogeny and evolution of the sound-generating structures in the infraorder Delphinida (Odontoceti: Delphinida). Biological Journal of the Linnean Society, 128(3), 700-724.
- Geisler JH, Colbert MW, Carew JL. 2014. A new fossil species supports an early origin for toothed whale echolocation. Nature 508: 383–386.
- Klima M. 1999. Development of the cetacean nasal skul. Berlin: Springer.
- McKenna, M. F., Cranford, T. W., Berta, A., & Pyenson, N. D. (2012). Morphology of the odontocete melon and its implications for acoustic function. Marine Mammal Science, 28(4), 690-713.
- Cranford TW, Amundin M, Norris KS. 1996. Functional morphology and homology in the odontocete nasal complex: implications for sound generation. Journal of Morphology 228: 223–285.
- Cranford, T. W. 1999. The sperm whale nose: Sexual selection on a grand scale? Marine Mammal Science 15:1133–1157.