Interactions multitrophiques et diversité des communautés
L'étude des interactions multitrophiques en relation avec la diversité des communautés fait appel à des disciplines aussi diverses que la mycologie, l'entomologie, la nématologie, la dynamique des populations et l'écologie théorique, pour appréhender le fonctionnement des écosystèmes. Cette nouvelle approche collaborative de l'écologie communautaire permet de mettre en évidence l’incidence des modes d’interaction et des stratégies biologiques des espèces sur la diversité taxonomique et fonctionnelle locale.
Les écosystèmes sont caractérisés par la diversité d’espèces qu’ils présentent et, par corrélation, par la diversité des interactions entre ces espèces. Toutefois, l'écologie des communautés (e.g. ensemble de populations vivant à un endroit donné et à un moment donné) s’est longtemps attachée aux interactions entre deux niveaux trophiques (e.g. rang qu’occupe un organisme au sein d’une chaîne trophique), en particulier à celles entre les plantes et les herbivores et celles entre les proies et les prédateurs.
L’intérêt des écologistes pour des interactions plus complexes impliquant deux (ou plus) niveaux trophiques n’est que très récent (années 1980). Pourtant, il est désormais évident qu'une approche des interactions multitrophiques traduit de manière beaucoup plus réaliste la complexité des mécanismes et des forces conductrices d’un écosystème que ne le fait une approche plus simple[1].
Histoire des concepts: interactions multitrophiques et diversité des communautés
Dès 1958, le biologiste Charles Elton a mis en évidence les interactions trophiques directes. Ce n’est qu’à partir des années 1980, que les interactions trophiques complexes commencent à être étudiées. Le besoin d'intégrer la résistance des plantes hôtes et le contrôle biologique à la gestion des ravageurs afin de comprendre l’importance des interactions directes et indirectes dans les communautés écologiques, a alors conduit à un intérêt croissant pour les interactions tritrophiques entre les plantes, les herbivores et leurs ennemis naturels[1] - [2].
La première approche de la diversité des communautés est développée à partir des années 1960 et met en exergue le rôle de la compétition et de la facilitation dans le maintien de la stabilité des communautés. Dans cette optique, plus une communauté comprend d’espèces, plus elle présente d’interactions et moins le système est sensible à la perte d’une interaction entre deux espèces.
Au début des années 1970, une théorie inverse est proposée. Elle suppose que plus un système est complexe, moins il est stable. La multiplication des connections entre espèces impliquerait une propagation des perturbations plus forte et plus durable dans l’écosystème. Ce dernier aurait alors plus de difficulté à recouvrer son état d’équilibre en cas de dérèglement.
Dans les années 1990, de nouvelles avancées vont être faites dans l’étude des écosystèmes et des interactions au sein des communautés. Elles vont démontrer l’existence d’un lien étroit entre la variabilité des espèces et la fluctuation de l’environnement. Dans cette perspective, la stabilité de l’écosystème serait due à la différence entre les niches écologiques des différentes espèces ; la diversité spécifique aurait un réel potentiel stabilisateur sur les propriétés des écosystèmes. Selon la définition moderne de Hutchinson, la niche écologique se définit comme l’ensemble des conditions physico-chimiques et biologiques; incluant les ressources limitantes ; nécessaire à une espèce pour maintenir une population stable ou croissante.
En 2001, S. Hubbell, à travers la théorie neutre remet en question les approches de la diversité des communautés en supposant que toutes les espèces d’un même niveau trophique occupent la même niche écologique et que leurs probabilités de survie, de mortalité, de reproduction et de migration sont les mêmes[3]. La dynamique locale d’un assemblage d’espèces serait due à la stochasticité démographique et à la capacité d’immigration limitée des organismes.
Mesures des deux concepts
La diversité est un paramètre déterminant du fonctionnement des écosystèmes. De même, les interactions multitrophiques jouent un rôle clé dans la structuration des écosystèmes et régissent la dynamique des populations qui les composent. Ainsi, afin de comprendre le lien entre la diversité et les interactions multitrophiques plusieurs mesures peuvent être réalisées.
Diversité des communautés
Afin de caractériser la diversité on s’intéresse à la richesse spécifique qui est le nombre d’espèces présentes dans la communauté considérée.
La diversité spécifique peut se mesurer à différents niveaux :
- 1 : La diversité alpha (Dα) correspond à la diversité locale dans une communauté. Pour la quantifier on utilise l’indice de Shannon. Elle permet de rendre compte du nombre d’espèces du milieu considéré et de la répartition des individus au sein de ces espèces

- H': indice de biodiversité de Shannon
- i : une espèce du milieu d’étude
- S : richesse spécifique
- où pi : Proportion d’une espèce i par rapport au nombre total d’espèces (S) dans le milieu d’étude (ou richesse spécifique du milieu), qui se calcule de la façon suivante : p(i)=ni/N où ni est le nombre d'individus pour l'espèce i et N est l'effectif total (les individus de toutes les espèces).
- 2 : La diversité bêta (Dβ) est une mesure de la diversité entre communautés. Elle correspond au taux de remplacement des espèces le long d'un gradient (par exemple climatique, spatial, d'altitude), au sein d'une région donnée. On utilise des indices de similitudes pour la calculer tel que l’indice de Jaccard :
Soit deux ensembles A et B

Il varie entre 0 (aucune espèces communes) et 1 (toutes les espèces sont communes).
- 3 : La diversité gamma (Dγ) est la diversité régionale, résultant de la combinaison des diversités alpha et bêta. Elle traduit le taux d'addition de nouvelles espèces quand on échantillonne le même habitat en différents endroits. Lande propose une approche statistique permettant de partitionner la diversité gamma comme:
β

Interactions multitrophiques
La complexité d’un réseau d’interaction peut être quantifiée à l’aide de trois mesures de la structure du réseau[4].
- 1 : La connectance : c’est le nombre d’interactions réalisées entre les niveaux trophiques spécifiés (L) divisé par le nombre d’interactions potentielles (Lmax)
- 2 : La diversité des interactions : on le calcul en utilisant l’indice de Shannon mais en l'appliquant aux interactions


où pi est la proportion d'interaction i par rapport à la somme totale de n interactions observées.
- 3 : La force des interactions : pour cela, on calcule la moyenne des forces d’interactions par la moyenne arithmétique de toutes les interactions significatives

où pi est la proportion d'interaction i par rapport à la somme totale de n interactions observées.
L’intérêt croissant pour l’étude de la biodiversité et la prise de conscience des enjeux que représente l’accélération de son érosion ont conduit à l’intégration de nouveaux paramètres dans son évaluation. Ainsi, la mesure de la biodiversité va désormais bien au-delà de l’estimation du nombre d’espèces dans un milieu mais prend en considération des facteurs comme la diversité fonctionnelle ou phylogénétique.
Interactions multitrophiques et biodiversité
Les interactions multitrophiques entre espèces et leurs conséquences sur la diversité des communautés sont généralement peu visibles dans des conditions stables et elles ne sont souvent mises en évidence qu'après la perte d’une ou plusieurs espèces. De plus, les échelles d’analyse des effets de cascade trophique sont souvent beaucoup plus vastes que celles prises en compte par la plupart des études scientifique.
Appréhender les liens entre les interactions multitrophiques et la biodiversité d’un milieu implique donc une approche pluriscalaire, visant à analyser le réseau et ses sous-ensembles. En effet, si on considère le gradient latitudinal, les variations des conditions abiotiques telles que la température, l’humidité et les capacités de dispersion et de colonisation des espèces sont prédominantes. A l’échelle locale, ce sont les processus biotiques (prédation, compétition et mutualisme) qui sont les plus impactant. L’échelle temporelle doit également être prise en compte pour analyser les interactions.
Diversité des interactions et stabilité des écosystèmes
Des modèles analytiques ont démontré que la multiplicité des interactions serait un indicateur de diversité et de stabilité de l’écosystème[5]. Ainsi, dans le cas d’une espèce généraliste (e.g. se nourrissant de plusieurs ressources), l’espèce pourra survivre en cas de disparition d’une de ses ressources car elle pourra se concentrer sur les autres. Dans cet exemple, la pluralité des interactions permet une certaine stabilité mais cette configuration n’est pas systématique car il est nécessaire de prendre en compte des variables environnementales. Les interactions au sein des écosystèmes se caractérisent par des flux de nature énergétique (carbone par exemple) le long des chaînes trophiques.
L’étude des relations entre les milieux dits aériens et souterrains met également en évidence l’importance des interactions dans le fonctionnement des écosystèmes. Dans ce contexte, les plantes jouent un rôle majeur car elles sont les médiateurs entre les organismes des deux milieux[6]. Les organismes structurant les réseaux trophiques sont de diverses natures : herbivores, carnivores, végétaux, champignons, bactéries, parasites et on estime que de très nombreux organismes participent indirectement à ces interactions mais demeurent non-détectés . On considère que la diversité végétale augmente les interactions et leurs forces respectives, modifiant ainsi la complexité du réseau [4]. Néanmoins, les effets de la diversité végétale s’atténuent avec l’augmentation du niveau trophique : les prédateurs sont moins influencés par la variation de cette diversité[7]. Aussi, les mycorhizes peuvent influencer la composition spécifique des plantes. Les micro-organismes interviennent dans les chaînes trophiques par des mécanismes pathogènes, par des mécanismes visant à modifier les qualités nutritionnelles des plantes ou par des mécanismes symbiotiques.
Le mutualisme peut alors être source de maintien de la diversité spécifique dans une communauté. En effet il peut permettre d’augmenter les capacités compétitrices d’une espèce, optimisant ainsi son développement. Dans le cas des symbioses mycorhiziennes, des plantes originellement peu compétitrices peuvent capter les ressources du sol plus facilement grâce aux hyphes (éléments filamenteux des champignons), améliorant leur croissance dans un milieu où elles se seraient faites exclure compétitivement par d’autres espèces en l’absence de mycorhizes. A travers la symbiose mycorhizienne, le mutualisme permet ici le maintien d’une diversité spécifique[1].
Ainsi, le maintien de cette diversité spécifique s’organise par des processus écologiques permettant la promiscuité entre espèces. Les processus locaux tel que la compétition, la prédation, le mutualisme permettent l’acquisition de ressources, tandis que les processus régionaux comme la dispersion permettent l’acquisition de territoires. Dans ce contexte, la cohabitation entre deux espèces repose sur un trade-off, c’est-à-dire un compromis entre bonne colonisation et bonne assimilation de ressources. (table1)

Si le rôle des interactions horizontales (au sein d’un même niveau trophique) sur le fonctionnement des écosystèmes semble prédictible, les interactions dites verticales, au sein de plusieurs niveaux trophiques semblent plus complexes.
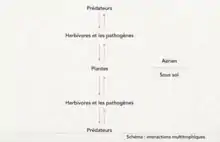
On distingue différentes dynamiques d’interactions verticales[8] :
- le contrôle « top-down » correspond à un mécanisme partant d’un niveau élevé d’une chaîne trophique à un niveau inférieur. C’est un contrôle descendant qui s’intéresse aux patrons généraux. Cette forme d’interaction présente une covariance négative, c’est-à-dire que l’on prévoit une quantité de biomasse variable selon les niveaux.
- le contrôle « bottom-up », à l’inverse, correspond à un mécanisme initié dans un niveau inférieur d’une chaîne trophique et qui influence les niveaux supérieurs. C’est un contrôle ascendant, qui s’intéresse plutôt à la compréhension des processus d’interaction à échelle réduite. Ici, la covariance est positive, c’est-à-dire que l’on prévoit une augmentation de la biomasse. (Schéma)
Espèces clés de voûte et biodiversité
Les études des interactions multitrophiques se concentrent souvent sur de petites zones et sur des espèces non mobiles, avec des cycles de vie courts, ce qui les rend faciles à observer. Il en résulte en une image incomplète et déformée du rôle des espèces clé de voûte dans les milieux naturels. Lorsque les populations de ces top-consommateurs sont réduites ou supprimées, et que les échelles spatiales et temporelles considérées sont suffisamment grandes, leur influence devient évidente.
Le lien entre les « apex-consommateurs » et la diversité des espèces peut se faire par l’intermédiaire de différentes interactions. Par exemple, en bloquant l'exclusion compétitive ou la prolifération des prédateurs intermédiaires (mesopredator release), ou encore par des effets indirects sur l'habitat[9].
L'altération de la dynamique de recrutement des arbres et la transformation progressive des forêts en landes et prairies en raison de l'augmentation de l'herbivorie des ongulés illustre l'influence des espèces clé de voûte sur la diversité fonctionnelle. Ce processus a été particulièrement mis en évidence dans les parcs naturels nord-américains où la perte de grands prédateurs au début du XXe siècle s’est traduite par une réduction du taux de croissance des arbres et par un échec du recrutement chez les essences dominantes. La réintroduction du loup au milieu des années 1990, dans le parc national de Yellowstone, a permis de réguler les populations de grands herbivores, de freiner la prolifération de coyotes, de restaurer la couverture végétale en diminuant la pression exercée par les grands ongulés et de stabiliser les berges des cours d’eau. Ce frein à l'érosion des berges et l’augmentation du refroidissement des rivières par l’ombre portée des arbres a permis une augmentation des espèces de poissons et d’autres organismes aquatiques[9]. L’installation des castors a également été favorisée et leurs barrages ont permis la création de niches écologiques propices aux loutres, ragondins, amphibiens, etc.
Dans un écosystème où l’influence du contrôle top-down est prépondérante, la perte de l’espèce située au sommet de la chaîne trophique engendre des conséquences sur les écosystèmes qui vont bien au-delà de la seule diversité de la communauté : augmentation des maladies et des incendies de forêt, pertes de séquestration du carbone, apparition d'espèces envahissantes, perturbation des cycles biogéochimiques. Ces impacts sont d’autant plus prégnants qu’ils sont exacerbés par des facteurs tels que la surexploitation des terres arables et des ressources, le changement climatique, la perte et la fragmentation des habitats, la pollution, etc[9] - [10].
Notes et références
- (en) Teja Tscharntke et Bradford A Hawkins, Multitrophic level interactions, Cambridge University Press, , 284 p. (ISBN 978-0-521-79110-6, OCLC 70747276, lire en ligne)
- (en) Wim H. Van der Putten, Louise E.M. Vet, Jeffrey A. Harvey et Felix L. Wäckers, « Linking above- and belowground multitrophic interactions of plants, herbivores, pathogens, and their antagonists », Trends in Ecology & Evolution, vol. 16, no 10, , p. 547–554 (DOI 10.1016/s0169-5347(01)02265-0, lire en ligne, consulté le )
- J.M. Levine J. HilleRisLambers, « The importance of niches for the maintenance of species diversity », Nature, vol 461, , p. 254-257 (DOI doi:10.1038/nature08251, lire en ligne)
- (en) Michael Rzanny et Winfried Voigt, « Complexity of multitrophic interactions in a grassland ecosystem depends on plant species diversity », Journal of Animal Ecology, vol. 81, no 3, , p. 614–627 (ISSN 1365-2656, DOI 10.1111/j.1365-2656.2012.01951.x, lire en ligne, consulté le )
- (en) A. Mougi et M. Kondoh, « Diversity of Interaction Types and Ecological Community Stability », Science, vol. 337, no 6092, , p. 349–351 (ISSN 0036-8075 et 1095-9203, PMID 22822151, DOI 10.1126/science.1220529, lire en ligne, consulté le )
- (en) Nicole M. van Dam et Martin Heil, « Multitrophic interactions below and above ground: en route to the next level », Journal of Ecology, vol. 99, no 1, , p. 77–88 (ISSN 1365-2745, DOI 10.1111/j.1365-2745.2010.01761.x, lire en ligne, consulté le )
- (en) Christoph Scherber, Nico Eisenhauer, Wolfgang W. Weisser et Bernhard Schmid, « Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment », Nature, vol. 468, no 7323, , p. 553–556 (ISSN 1476-4687, DOI 10.1038/nature09492, lire en ligne, consulté le )
- (en) Andreas Schuldt and al, « Belowground top-down and aboveground bottom-up effects structure multitrophic communitiy relationships in a biodiverse forest », Nature, vol. 7, no 4222, (DOI 10.1038/s41598-017-04619-3, lire en ligne)
- (en) Estes J. A an al, « Trophic downgrading of planet Earth », Science, , p. 301-306 (DOI 10.1126/science.1205106, lire en ligne)
- (en) Johnson, M.T.J, Marc J. Lajeunesse et Anurag A. Agrawal, « Additive and interactive effects of plant genotypic diversity on arthropod communities and plant fitness », Ecology letters, , p. 24-34 (DOI 10.1111/j.1461-0248.2005.00833.x, lire en ligne)