Coude (biochimie)
En biochimie, un coude est un élément de la structure secondaire des protéines caractérisé par un changement de direction de la chaîne polypeptidique. On peut définir un coude comme une configuration tridimensionnelle dans laquelle les atomes de carbone α de deux résidus d'acides aminés séparés par un petit nombre d'autres résidus — typiquement moins de cinq — sont distants de moins de 0,7 nm[1] en dehors d'un élément de structure secondaire tels qu'une hélice α ou un feuillet β avec un squelette peptidique formant une succession d'angles dièdres régulière[2]. La présence d'une liaison hydrogène dans une telle structure est fréquemment observée mais n'est pas une condition nécessaire pour qu'on soit en présence d'un coude.
Typologie
On classe généralement les coudes en fonction du nombre de résidus qu'ils comprennent[3] :
- coude α : 4 liaisons peptidiques, et donc 3 résidus, séparent les deux résidus extrêmes de la structure[4] ;
- coude β : 3 liaisons peptidiques, et donc 2 résidus, séparent les deux résidus extrêmes de la structure ; c'est le type de coude le plus fréquent dans les protéines ;
- coude γ : 2 liaisons peptidiques, et donc 1 résidus, séparent les deux résidus extrêmes de la structure[5] ;
- coude δ : les deux résidus extrêmes de la structure son contigus, c'est-à-dire qu'ils ne sont séparés que par une seule liaison peptidique (structure peu probable d'un point vue stérique) ;
- coude π : 5 liaisons peptidiques, et donc 4 résidus, séparent les deux résidus extrêmes de la structure[6].
On parle parfois de boucle ω pour qualifier les structures contenant davantage de résidus au sein desquelles les liaisons hydrogène ne sont pas fixes.
Il est fréquent qu'un ou plusieurs résidus soient impliqués dans deux coudes qui se recouvrent partiellement. C'est par exemple le cas d'une séquence de cinq résidus dans laquelle les résidus 1-4 et 2-5 forment des coudes : on parle dans ce cas de coude double (I, I+1). Les coudes multiples (jusqu'à sept fois) sont des structures courantes dans les protéines[7].
Les épingles à cheveux sont un cas particulier de coude dans lequel le sens du squelette peptidique s'inverse et interagit avec les structures secondaires voisines. Par exemple, une épingle à cheveux β relie deux brins β antiparallèles avec des liaisons hydrogène[8].
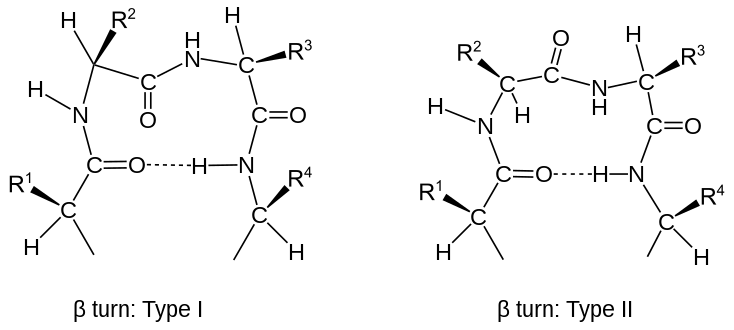 (en) Coude β de type I à gauche et de type II à droite.
(en) Coude β de type I à gauche et de type II à droite.
Chaque type de coude peut être subdivisé en fonction de l'angle dièdre de son squelette carboné (diagramme de Ramachandran). Ainsi, changer le signe de ses angles dièdres conduit à inverser la chiralité de la chaîne peptidique — on n'obtient cependant pas l'énantiomère de la protéine car la chiralité de chaque atome de carbone α est conservée. Il existe ainsi deux formes de coude γ : une forme classique dont les angles dièdres (φ, ψ) sont à peu près (75°, –65°), et une forme inverse ayant pour angles dièdres (–75°, 65°). Il existe au moins huit formes de coude β, en fonction de la présence d'un isomère cis de la liaison peptidique et des angles dièdres deux deux résidus encadrés par les deux résidus refermant ce type de coude. On distingue les coudes β classiques et inverses par un signe prime : ainsi un coude β de type I’ est inverse d'un coude β de type I. Si l'on définit les résidus extrêmes encadrant un coude par la présence d'une liaison hydrogène entre eux, les quatre catégories I, I’, II’ et II de Venkatachalam[9] sont suffisantes pour décrire tous les coudes β possibles[10]. Tous les quatre sont fréquents dans les protéines, le coude β de type I étant le plus fréquent, suivi par les coudes β de types II, I’ et II’ respectivement.
Rôle dans le repliement des protéines
Deux approches opposées ont été proposées pour décrire le rôle des coudes dans le repliement des protéines. La première les considère comme des éléments passifs qui jouent simplement le rôle de connecteurs entre éléments actifs de la structure tridimensionnelle de la protéine. Cette vision est confortée par le fait que la séquence est généralement très peu conservée au niveau des coudes. A contrario, les coudes peuvent être vus comme des éléments actifs qui jouent un rôle important en permettant aux autres éléments de la structure secondaire des protéines de s'agencer dans l'espace conformément à la forme native de la protéine. Cette vision est confortée par le fait que certaines mutations au niveau des coudes empêchent certaines protéines de se replier de manière fonctionnelle.
Notes et références
- (en) C. M. Wilmot et J. M. Thornton, « Analysis and prediction of the different types of β-turn in proteins », Journal of Molecular Biology, vol. 203, no 1, , p. 221-232 (PMID 3184187, DOI 10.1016/0022-2836(88)90103-9, lire en ligne)
- (en) George D. Rose, Lila M. Glerasch et John A. Smith, « Turns in Peptides and Proteins », Advances in Protein Chemistry, vol. 37, , p. 1-109 (PMID 2865874, DOI 10.1016/S0065-3233(08)60063-7, lire en ligne)
- (en) Claudio Toniolo et Ettore Benedetti, « Intramolecularly Hydrogen-Bonded Peptide Conformation », Critical Reviews in Biochemistry, vol. 9, no 1, , p. 1-44 (PMID 6254725, DOI 10.3109/10409238009105471, lire en ligne)
- (en) Vicenzo Pavone, Girolamo Gaeta, Angela Lombardi, Flavia Nastri, Ornella Maglio, Carla Isernia et Michele Saviano, « Discovering protein secondary structures: Classification and description of isolated α-turns », Biopolymers, vol. 38, no 6, , p. 705-721 (PMID 8652792, DOI 10.1002/(SICI)1097-0282(199606)38:6<705::AID-BIP3>3.0.CO;2-V, lire en ligne)
- (en) E. James Milner-White, « Situations of gamma-turns in proteins: Their relation to alpha-helices, beta-sheets and ligand binding sites », Journal of Molecular Biology, vol. 216, no 2, , p. 385-397 (PMID 2254936, DOI 10.1016/S0022-2836(05)80329-8, lire en ligne)
- (en) K. R. Rajashankar et S. Ramakumar, « π-Turns in proteins and peptides: Classification, conformation, occurrence, hydration and sequence », Protein Sciences, vol. 5, no 5, , p. 932-946 (PMID 8732765, PMCID 2143406, DOI 10.1002/pro.5560050515, lire en ligne)
- (en) E. Gail Hutchinson et Janet M. Thornton, « A revised set of potentials for β-turn formation in proteins », Protein Science, vol. 3, no 12, , p. 2207-2216 (PMID 7756980, PMCID 2142776, DOI 10.1002/pro.5560031206, lire en ligne)
- (en) Bancinyane Lynn Sibanda, Tom L. Blundell et Janet M. Thornton, « Conformation of β-hairpins in protein structures: A systematic classification with applications to modelling by homology, electron density fitting and protein engineering », Journal of Molecular Biology, vol. 206, no 4, , p. 759-777 (PMID 2500530, DOI 10.1016/0022-2836(89)90583-4, lire en ligne)
- (en) C. M. Venkatachalam, « Stereochemical criteria for polypeptides and proteins. V. Conformation of a system of three linked peptide units », Biopolymers, vol. 6, no 10, , p. 1425-1436 (PMID 5685102, DOI 10.1002/bip.1968.360061006, lire en ligne)
- (en) Jane Shelby Richardson, « The Anatomy and Taxonomy of Protein Structure », Advances in Protein Chemistry, vol. 34, , p. 167-339 (PMID 7020376, DOI 10.1016/S0065-3233(08)60520-3, lire en ligne)