Cortex moteur
Le cortex moteur désigne l'ensemble des aires du cortex cérébral qui participent à la planification, au contrôle et à l'exécution des mouvements volontaires des muscles du corps. D'un point de vue anatomique, le cortex moteur est situé dans la partie postérieure du lobe frontal, au niveau de la région caudale de la circonvolution frontale ascendante en avant du sillon central. Le cortex moteur est en interaction constante avec d'autres structures nerveuses impliquées dans le mouvement comme le système des ganglions de la base et le cervelet.
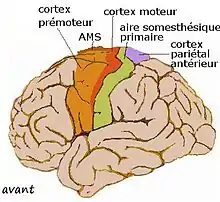
Divisions du cortex moteur
Le cortex moteur peut être divisées en plusieurs aires fonctionnelles :
- le cortex moteur primaire ou aire motrice primaire (noté M1) commande directement l'exécution du mouvement ;
- le cortex moteur secondaire (M2) ou cortex pré-moteur inclut :
- le cortex prémoteur latéral ou aire prémotrice situé sur la surface externe du lobe frontal, en avant de M1 qui contrôle l'organisation des mouvements des muscles proximaux et de ceux du tronc,
- l'aire motrice supplémentaire (AMS) qui coordonne et planifie les gestes complexes impliquant une séquence de mouvements ou la coordination de plusieurs membres. L'AMS est située en avant du cortex moteur, sur la face interne des deux hémisphères,
- bien qu'il ne s'agisse pas d'un territoire du lobe frontal, on y adjoint parfois le cortex pariétal postérieur qui est impliqué dans la transformation des informations visuelles en informations spatiales, indispensables au guidage des mouvements.
Cortex moteur primaire
L'aire M1 correspond à l'aire 4 de Brodmann, située sur la paroi antérieure du sillon central. Sur le plan cytoarchitectonique, il s'agit d'un cortex hétérotypique agranulaire à six couches. C'est la cinquième couche (dite couche pyramidale interne) qui abrite les neurones pyramidaux géants (ou cellules de Betz) dont la taille atteint quelques dixièmes de millimètre et qui projettent leurs longs axones sur les motoneurones et les interneurones, formant le faisceau corticospinal de la moelle spinale
Aire motrice supplémentaire
L'aire motrice supplémentaire et l'aire prémotrice permettent la préparation du mouvement en fonction des stimuli externes (sensitive) ou internes (mémoire) reçus.
Cortex pariétal postérieur
Le cortex pariétal postérieur reçoit des informations somatosensorielles, proprioceptives et visuelles qui permettent d'évaluer le contexte. Il est étroitement interconnecté au cortex préfrontal.
Évolution du cortex moteur
Les mammifères ont évolué à partir de reptiles-mammifères il y a plus de 200 millions d'années[1]. Ces premiers mammifères ont développé plusieurs fonctions cérébrales nouvelles probablement en raison de nouveaux processus sensoriels nécessaires à la niche nocturne que ces mammifères occupaient[2]. Ces animaux longtemps recherchés possédaient probablement un cortex « somato-moteur », où l'information somato-sensorielle et l'information motrice étaient traitées dans la même région corticale. Cela leur a permis d'acquérir des compétences motrices simples, comme la locomotion quadrupède ou l'attaque et la fuite des prédateurs et des proies. Les mammifères placentaires ont développé un cortex moteur discret il y a environ 100 million d'années[1]. La masse d'un tissu nerveux contrôlant une fonction particulière étant proportionnelle à la quantité de traitement de l'information impliquée dans l'exécution de la fonction[2], les mammifères placentaires ont développé un cortex moteur séparé. Ces mammifères et les compétences motrices acquises par ces organismes étaient plus complexes que leurs premiers ancêtres mammifères. En outre, ce cortex moteur a permis le développement des modes de vie arboricoles de nos ancêtres primates.
Les améliorations apportées au cortex moteur et la présence de pouces opposables et de vision stéréoscopique ont été privilégiées de manière évolutive pour empêcher les primates de se tromper lors des sauts entre les branches d'arbres[3]. En raison de cette pression évolutive, le système moteur des primates arborescents présente un degré disproportionné de représentation somatotopique des mains et des pieds, qui sont essentiels à la saisie des branches[4] - [5] - [6].
Notes et références
Références
- Jon H. Kaas, « Evolution of somatosensory and motor cortex in primates », The Anatomical Record. Part A, Discoveries in Molecular, Cellular, and Evolutionary Biology, vol. 281, no 1, , p. 1148–1156 (ISSN 1552-4884, PMID 15470673, DOI 10.1002/ar.a.20120, lire en ligne, consulté le )
- (en) Harry Jerison, Evolution of The Brain and Intelligence, Elsevier, , 496 p. (ISBN 978-0-323-14108-6, lire en ligne)
- (en) Farish A. Jr Jenkins, Primate Locomotion, Elsevier, , 402 p. (ISBN 978-0-323-15202-0, lire en ligne)
- Atsushi Nambu, « Somatotopic Organization of the Primate Basal Ganglia », Frontiers in Neuroanatomy, vol. 5, (ISSN 1662-5129, PMID 21541304, PMCID PMC3082737, DOI 10.3389/fnana.2011.00026, lire en ligne, consulté le )
- T. P. Pons, P. E. Garraghty, C. G. Cusick et J. H. Kaas, « The somatotopic organization of area 2 in macaque monkeys », The Journal of Comparative Neurology, vol. 241, no 4, , p. 445–466 (ISSN 0021-9967, PMID 4078042, DOI 10.1002/cne.902410405, lire en ligne, consulté le )
- (en) G. Rizzolatti, R. Camarda, L. Fogassi et M. Gentilucci, « Functional organization of inferior area 6 in the macaque monkey », Experimental Brain Research, vol. 71, no 3, , p. 491–507 (ISSN 0014-4819 et 1432-1106, DOI 10.1007/BF00248742, lire en ligne, consulté le )