ADN satellite
L'ADN satellite est un type de séquences d’ADN répétées.
Composition
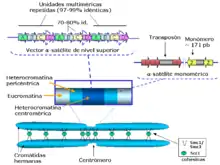
L’ADN satellite se présente sous la forme de séquences d’ADN pouvant atteindre plusieurs millions de paires de bases est constitué d’un grand nombre de répétitions d’une séquence beaucoup plus petite (l’unité de répétition). La longueur de l’unité de répétition est variable d’un type d’ADN satellite à l’autre, de 5 paires de bases pour les satellites III humains à 755 paires de bases pour les satellites trouvés dans les génomes des plantes[1].
Le nom « ADN satellite » vient de ce que l’ADN de ces séquences, en raison de leur nature répétitive, a une composition en nucléotides fortement biaisée par rapport au reste du génome ; en conséquence, il sédimente différemment par ultracentrifugation (accumulation dans le tube de centrifugation à un endroit qui dépend précisément de sa densité) et forme une bande secondaire, dite « satellite », dans un gradient de chlorure de césium[2].
Génome humain
Le génome humain comprend environ 7 % d’ADN satellite[1].
L’ADN α-satellite (aussi appelé alphoid DNA en anglais) est constitué de répétitions d’un motif de 171 paires de bases[3]. On le retrouve au niveau du centromère de tous les chromosomes, où il s’étend sur des régions allant de quelques centaines de kilobases à plusieurs mégabases — le nombre de répétitions et la taille totale étant variables d’un chromosome à l’autre et d’un individu à l’autre. L’ADN α-satellite contribue à la fonction du centromère, en permettant notamment le recrutement de la protéine CENP-B (Centromeric protein B) par l’intermédiaire d’un motif conservé de 17 paires de base. Néanmoins, il n’est pas indispensable à la bonne fonction du centromère, des néocentromères pleinement fonctionnels pouvant être observés sur des régions génomiques dépourvues d’α-satellites[4].
L’ADN β-satellite est constitué de répétitions d’un motif de 68 paires de bases[5]. Il s’étale sur des régions de 50 à 300 kilobases sur les bras courts des chromosomes acrocentriques (chromosomes 13, 14, 15, 21 et 22), sur les péricentromères des chromosomes 1, 3 et 9 ainsi que sur le bras long du chromosome Y[6].
L’ADN satellite III est constitué de répétitions d’un motif de 5 paires de bases (GGAAT), occupant plusieurs centaines de kilobases dans la région péricentromérique de la plupart des chromosomes[1].
Génome murin
L’ADN satellite représente environ 3,5 % du génome murin[7] et est associé aux régions centromériques et péricentromériques de tous les chromosomes. Il en existe de deux types :
Notes et références
- Richard, Kerrest et Dujon 2008
- (en) SaulKit, « Equilibrium sedimentation in density gradients of DNA preparations from animal tissues », Journal of Molecular Biology, vol. 3, no 6, , p. 711–716 (DOI 10.1016/S0022-2836(61)80075-2).
- Sullivan, Schwartz et Willard 1996
- Karpen et Allshire 1997
- Waye et Willard 1989
- Meneveri et al. 1993
- Mouse Genome Sequencing Consortium 2002
- Hörz et Altenburger 1981
- Wong et Rattner 1988
Voir aussi
Bibliographie
- (en) Wolfram Hörz et Werner Altenburger, « Nucleotide sequence of mouse satellite DNA », Nucleic Acids Res., vol. 9, no 3, , p. 683–696 (PMID 6261227, lire en ligne [PDF])
- (en) Gary H. Karpen et Robin C. Allshire, « The case for epigenetic effects on centromere identity and function », Trends Genet., vol. 13, no 12, , p. 489–496 (PMID 9433139)
- (en) R. Meneveri, A. Agresti, A. Marozzi, S. Saccone, M. Rocchi, N. Archidiacono, G. Corneo, G. Della Valle et E. Ginelli, « Molecular organization and chromosomal location of human GC-rich heterochromatic blocks », Gene, vol. 123, no 2, , p. 227–234 (PMID 8428662)
- (en) Mouse Genome Sequencing Consortium, Robert Waterston, Kerstin Lindblad-Toh, Ewan Birney, Jane Rogers, Josep Abril, Pankaj Agarwal, Richa Agarwala, Rachel Ainscough, Marina Alexandersson, Peter An, Stylianos Antonarakis, John Attwood, Robert Baertsch et Jonathon Bailey, « Initial sequencing and comparative analysis of the mouse genome », Nature, vol. 420, no 6915, , p. 520–562 (PMID 12466850, DOI 10.1038/nature01262)
- (en) Guy-Franck Richard, Alix Kerrest et Bernard Dujon, « Comparative genomics and molecular dynamics of DNA repeats in eukaryotes », Microbiol. Mol. Biol. Rev., vol. 72, no 4, , p. 686–727 (PMID 19052325, DOI 10.1128/MMBR.00011-08, lire en ligne [PDF])
- (en) Beth A. Sullivan, Stuart Schwartz et Huntington F. Willard, « Centromeres of human chromosomes », Environ. Mol. Mutagen., vol. 28, no 3, , p. 182–191 (PMID 8908179, DOI 10.1002/(SICI)1098-2280(1996)28:3<182::AID-EM4>3.0.CO;2-G)
- (en) John S. Waye et Huntington F. Willard, « Human beta satellite DNA: genomic organization and sequence definition of a class of highly repetitive tandem DNA », Proc. Natl. Acad. Sci. U.S.A., vol. 86, no 16, , p. 6250–6254 (PMID 2762326, lire en ligne [PDF])
- (en) A. K. C. Wong et J. B. Rattner, « Sequence organization and cytological localization of the minor satellite of mouse », Nucleic Acids Res., vol. 16, no 24, , p. 11645–11661 (PMID 3211746, lire en ligne [PDF])