Théorie métabolique de l'écologie
La théorie métabolique de l'écologie (MTE, pour Metabolic theory of ecology en anglais), est une théorie quantitative qui permet d'expliquer le lien entre le métabolisme, la taille des individus et leur température.
Elle fut introduite par Arrhenius et Kleiber et synthétisée en 2004 dans un article rédigé par J.H Brown et al., "Toward a metabolic theory of ecology"[1].
Cette théorie prédit comment le métabolisme contrôle les processus écologiques à l'échelle des individus, des populations et des écosystèmes.
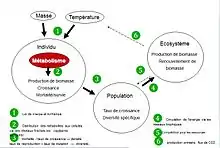
Le métabolisme d'un organisme correspond à l'ensemble des transformations biochimiques qui se produisent à l'intérieur de son corps. Les organismes prélèvent l'énergie nécessaire à leur métabolisme dans leur environnement. Les besoins énergétiques varient d'un organisme à l'autre, en fonction de leur taille et leur température corporelle.
La théorie métabolique, bien que reprise par de nombreux travaux en écologie, rencontre actuellement des limites, et est fortement controversée.
Les travaux de Kleiber et Arrhenius
Des progrès récents ont permis de comprendre le rôle de la température, de la taille du corps et la de stœchiométrie dans les mécanismes biologiques. Ces paramètres agissant à des niveaux d’organisation moléculaire, cellulaire et à celui de l’organisme tout entier, ont permis de fonder une théorie métabolique de l’écologie.
La théorie métabolique relie la performance individuelle des organismes à l’écologie des populations, des communautés et des écosystèmes.
Le métabolisme est un processus biologique unique qui obéit à des principes physiques et chimiques qui gouvernent la transformation d’énergie et de matière. Il détermine les demandes que les organismes placent dans leur environnement pour toutes les ressources et simultanément définit de puissantes contraintes sur l’allocation des ressources à toutes les composantes de la fitness.
Le taux métabolique est fondamental en biologie. Il représente la vitesse à laquelle les organismes absorbent, transforment et allouent l’énergie et la matière. Il est souvent mesuré en suivant la consommation d'O2. Le taux métabolique est gouverné par trois facteurs : la taille du corps, la température et la stœchiométrie.
Les caractéristiques des organismes varient de façon prévisible avec la taille du corps. Les travaux de Max Kleiber (1932) ont permis d'établir une loi scalante entre la masse des organismes et leur taux métabolique, pour une température corporelle donnée. La loi de Kleiber prédit que :
(1) Q = q.M3/4
Q: le taux métabolique, q: une constante indépendante de la taille du corps, M: la masse de l’organisme.
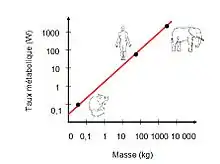
Plus un animal est grand, plus son métabolisme sera élevé : un éléphant consomme beaucoup plus d'O2 qu'une souris. En revanche, si l'on considère le métabolisme spécifique par unité de poids, on aura l'effet inverse:
(2) Q/M = q.(M3/4/M) = q.M-1/4 = B (kcal/jour/kg)
Un animal de grande taille a un métabolisme spécifique plus faible. Cette corrélation s'explique par le rapport surface/volume des organismes. Une souris possède une grande surface pour un petit volume. À l'inverse de l'éléphant, elle subit donc beaucoup plus de pertes de chaleur par diffusion. Par extension, beaucoup d'autres paramètres physiologiques suivent cette loi de M3/4[2] et ont une incidence sur divers paramètres des organismes.
La température augmente exponentiellement la vitesse des réactions biochimiques (notamment la cinétique des enzymes), les taux métaboliques ainsi que d'autres activités biologiques. Cette cinétique est représentée par la constante de Boltzmann dans la réaction de Van’t Hoff- Arrhenius :
(3) B = q.M-1/4.e -E/kT
E est l'énergie d’activation en électron volt, k: la constante de Boltzmann (elle montre comment la température affecte la vitesse des réactions en changeant la proportion des molécules avec une énergie cinétique suffisante) et T: la température en kelvin.
Cette équation peut être simplifiée:
(4) B ∝ M-1/4.e-E/kT
C'est l'équation établie par Brown et al. et mettant en évidence une relation entre métabolisme et masse, dépendante de la température.
Brown et al. prennent aussi en compte la relation établie par Gillooly et Allen qui concerne le taux de renouvellement tB des substrats énergétiques:
(5) tB ∝ M1/4.e E/kT
Cette relation illustre qu'un animal de grande taille consomme plus de ressources qu'un de petite taille, et qu'il les distribue plus lentement.
Les fondements mécanistiques de la théorie
Le modèle de West et Brown considère que l'activité métabolique des organismes (par exemple la respiration, la synthèse de nouveaux tissus, la production primaire) est limitée par le transport des nutriments et leur distribution dans le corps[3].
Chez les animaux et les plantes, le transport des nutriments s'effectue par des systèmes vasculaires. Pour les animaux, ce système consiste en un réseau de capillaires dont le diamètre diminue au fur et à mesure que le réseau s'étend. Ces réseaux de capillaires transportent également l'oxygène, accepteur d'électrons essentiels à un métabolisme aérobie.
L'efficacité de distribution des ressources (nutriments et oxygène) via ces réseaux contribuent au bon fonctionnement du métabolisme des organismes.
West et Brown considèrent que la structure de ces réseaux de capillaires est similaire chez tous les organismes, car ils suivent une loi allométrique universelle : la taille de ces réseaux est proportionnelle à la masse des organismes, élevée à la puissance ¾. De plus, ces réseaux suivent des lois fractales (ils ont la même géométrie à différentes échelles), et assurent l'approvisionnement en nutriments de toutes les cellules[4].
Ces conditions expliquent que l'activité métabolique soit aussi proportionnelle à la masse corporelle à la puissance ¾.
Une telle supposition équivaut à dire que l'évolution a convergé vers l'apparition de systèmes capillaires plus efficaces, en minimisant l'énergie et le temps alloué au transport de nutriments.
Les prédictions de la théorie
Les caractéristiques des individus
De nombreux taux biologiques sont dépendants de la masse et de la température suivant l'équation (4)
Production individuelle de biomasse
La production de biomasse d'un organisme suit une équation du type P ∝ M3/4.e-E/kT
On peut en déduire la production de biomasse spécifique: P/M ∝ M-1/4.e-E/kT
La production de biomasse spécifique P/M suit une équation semblable au taux métabolique spécifique, ces deux paramètres évoluent ensemble avec la température et la taille. D'après Brown ceci soutient l’hypothèse selon laquelle une part constante du métabolisme est allouée à la production de biomasse. La MTE prédit que deux organismes différents, de même masse et à la même température ont une production de biomasse similaire[1]. Il existe cependant une variation résiduelle entre les différents groupes, probablement liée à des facteurs spécifiques de l'espèce ou de l’environnement.
La réciprocité entre les temps métaboliques et le taux métabolique suit la relation : (5) tB ∝ M1/4.eE/kT[5]
La croissance de l'organisme dépend donc de la masse et de la température : plus l'organisme est gros et la température est basse, plus les temps métaboliques sont longs.
Survie et mortalité
Pour une population stable, la mortalité doit être égale à la fécondité. Cette dernière est liée à la production de biomasse et la MTE prédit que la mortalité (Z) dépend du taux métabolique spécifique : Z ∝ M-1/4.e-E/kT
D'après la MTE, plus l'organisme est grand et la température basse, plus sa mortalité sera faible[6] - [7].
La mortalité est influencée par la compétition, la prédation et le parasitisme. Ces interactions écologiques dépendent en partie de la température.
Taux de croissance des populations Rmax
La théorie métabolique de l'écologie montre qu'il est possible d'estimer le taux de croissance exponentielle Rmax grâce à la masse spécifique de l'organisme et la température.
L'âge de la première reproduction est corrélé positivement avec la masse de l'organisme. Des individus de petite taille se reproduisent plus rapidement.
D'après la théorie, il est possible de déterminer le taux de croissance d'une population grâce au métabolisme de l’espèce considérée. En microbiologie, par exemple, le taux métabolique d'une bactérie peut être déterminé grâce au taux de croissance maximal d'une population. Pour les eucaryotes, la théorie métabolique de l'écologie prévoit que Rmax∝ M-1/4 et que Rmax.M-1/4 ∝ 1/kT .
La théorie montre que la dynamique d'une population possède des contraintes allométriques et cinétiques liées au métabolisme. Par conséquent, les petits organismes qui ont une température interne élevée possèdent un Rmax supérieur aux organismes plus grands, avec une température interne plus froide.
Densité de population
L'équilibre du nombre d'individus ou de la capacité de charge K est prédit par une relation linéaire dépendante des ressources R, paramètre limitant de l'augmentation de la densité de la population:
(6) K∝ [R]M-3/4eE/kT
D'après cette équation, on peut dire que la population sera plus dense si on augmente les ressources, et si la taille des individus est petite. En revanche, la capacité de charge est réduite si la température augmente. Cette théorie a été testée chez les plantes et semble se révéler exacte[1]. Cependant, pour les animaux mobiles, d'autres paramètres entrent en jeu et le modèle est moins bien adapté. Il existe une très grande hétérogénéité des milieux environnementaux, des régimes trophiques et des modes de dispersion des populations, selon les espèces.
Interactions inter-spécifiques
De nombreuses expériences ont tenté de démontrer comment les proies, prédateurs et espèces compétitrices pouvaient coexister dans le même environnement. Il est difficile de démontrer la stabilité de ces interactions en laboratoire, elle conduit en général à l'extinction d'une des espèces étudiées. En modifiant la température, les résultats de la compétition peuvent être inversés : l'espèce gagnante devient perdante (Park, 1954). Ces résultats sont expliqués par la modification de l'optimum de l'espèce compétitrice gagnante lorsqu'il y a modification de la température. Le temps de l'exclusion compétitive varie avec la température. La MTE prédit les taux de déclin et de croissance des populations compétitrices qui sont déterminées par les taux individuels du métabolisme.
Diversité spécifique
Les modèles empiriques qualitatifs de la biodiversité suggèrent que les processus métaboliques génèrent et maintiennent la diversité des espèces, en suivant l'équation (4). Ainsi, les petits organismes ont une diversité plus grande dans les milieux à température élevée par rapport à ceux des milieux à température basse. Il semblerait que la diversité d’une espèce varie inversement avec sa taille, a priori le métabolisme joue un rôle central.
La diversité de la plupart des groupes taxonomiques est plus élevée dans les tropiques. Or, cela a toujours été attribué à une plus grande productivité ou à la réduction des saisons, plutôt qu’à l’effet cinétique des températures élevées.
Allen et al en 2002[8] ont démontré que la diversité des espèces, chez de nombreux groupes de plantes et d'animaux, à différentes latitudes mais température égale, suit la même relation de Boltzmann que le taux métabolique (3). Ceci s’applique non seulement aux gradients des latitudes, mais aussi le long des gradients d’altitudes.
Dans des environnements à forte biodiversité, tels que les forêts tropicales et les récifs coralliens, on peut s’attendre à des interactions qui mènent à une exclusion rapide. Ces processus évolutifs obéissent aux équations (4) et (5). Chez les petits organismes, ou des organismes qui habitent les zones tropicales, on observe une dynamique évolutive plus grande et plus rapide par rapport à ceux de grande taille ou provenant d’un climat froid.
J.F Gilloly et A.P Allen[9] ont démontré que l'équation (5) permet de prédire le taux d’évolution moléculaire pour une diversité de gènes et des génomes chez des ectothermes et des endothermes. Par ailleurs, Van Valen (1973) attribue l’origine et le maintien de la biodiversité en grande partie au phénomène de la Reine Rouge. Ceci laisse à supposer que ce phénomène se déroule selon l'équation (5). Autrement dit, plus il fait chaud ou plus l'organisme est petit, plus le métabolisme est rapide, et inversement.
Les processus écosystémiques
La théorie métabolique permet de prévoir explicitement la contribution des biotes aux cycles biogéochimiques. Plus clairement, l'équation (4) fournit la base pour prédire comment la taille et la température des organismes déterminent les valeurs des réserves, ainsi que les taux des flux à l’intérieur et entre les compartiments pour les producteurs primaires, les herbivores, les prédateurs et les détritivores.
Des écosystèmes avec la même quantité de ressources, et à température égale, comme une forêt et une prairie adjacente, auront des taux de production primaire très proches. Cependant, la biomasse totale sera plus élevée pour la forêt.
La productivité dans un écosystème à l'équilibre (où la production de biomasse est égale à sa consommation), peut être interprétée comme le remplacement des pertes individuelles, dues à la mortalité.
En écologie on l'exprime comme étant le ratio de la productivité totale de la population (Ptot ∝ M3/4.e-E/kT) par le stock de biomasse présent dans le milieu (W).
Sachant que W ∝ M, alors Ptot/W ∝ M-1/4.e-E/kT (9)
On notera que la productivité suit la même loi que la mortalité : elle sera maximisée pour des organismes de petite taille, et pour des températures chaudes. Par conséquent, le temps de renouvellement de cette biomasse, qui est l'inverse de la productivité, doit être proportionnel à M1/4. Plus un écosystème est constitué de grands organismes au métabolisme lent, plus le renouvellement de la biomasse est lent. Ce temps est proche de 22 ans pour une forêt tandis qu'il avoisine les 3 ans pour une prairie.
La théorie pourrait alors s'appliquer aux dynamiques des successions qui interviennent après une perturbation de l'écosystème, tels feux de forêt, déforestation. En effet, on pourrait s'attendre à des changements de productivité, de biomasse, qui s'opèrent lors du passage d'écosystèmes dominés par les herbacés à ceux dominés par les arbres.
Limites et controverses
.jpg.webp)
Depuis la formulation de la MTE, beaucoup de tests et d'expériences ont été faits afin de déterminer les limites de cette théorie[10]. Ces tests pratiqués montrent que certaines hypothèses de la MTE sont valables, mais que d’autres hypothèses fondamentales sont biologiquement incompatibles. Les auteurs précisent donc que la MTE doit être affinée et améliorée[11]. De plus, de nombreuses hypothèses de la MTE n’ont pas encore été testées.
1. Les prédictions réfutées par des approches expérimentales
Entre la publication de l'article de Brown et al. en 2004, qui pose les fondements de la théorie, et l'année 2012, 560 articles ont été écrits en relation avec cette théorie, qui demeure l'objet de nombreuses critiques, notamment sur les prédictions de la MTE à grande échelle.
Par exemple, une étude[12] de 2007 démontre qu'il n'existe pas de corrélation positive entre la température et la richesse spécifique, contrairement aux prédictions de la MTE.
D'autres expériences ont montré que l’effet de la température ne pouvait pas être décrit correctement par l’équation d’Arrhénius[13]. Par exemple, le taux d’ingestion des proies par un prédateur, en utilisant une échelle logarithmique, ne suit pas de relation linéaire avec la température, et diminue à des températures trop élevées. La loi d’Arrhénius utilisée par la MTE n’est donc valable que pour des gammes de températures réduites et ne permet pas d’avoir un modèle général[14].
2. Les protistes, des organismes invalidant la théorie
La MTE prédit une relation entre la masse et le métabolisme, avec un exposant ¾ qui serait expliquée par l’organisation des réseaux de capillaires permettant le transport des différents métabolites au sein de l’organisme. Cependant, la relation avec l’exposant ¾ se retrouve également chez les protistes, qui ne possèdent aucun réseau de transport puisqu’il s’agit d’organismes « simples » sans tissus spécialisés, le plus souvent unicellulaires. L’explication de la présence de l’exposant ¾, sur laquelle repose toute la théorie métabolique de l’écologie, est donc invalidée par ce groupe.
3. La DBE, une théorie métabolique alternative
D’autres théories visant à comprendre les phénomènes métaboliques ont été proposées. Parmi celles-ci on trouve la théorie appelée « Dynamic Energy Budget », ou DEB. Les organismes sont décrits en termes de « Structure » et « Réserve ». La DEB cherche à déterminer les flux d’énergie et de masse entre ces compartiments de Structure et Réserve, et à modéliser les allocations d’énergie à différents paramètres tels que : reproduction, croissance, acquisition de la maturité, maintenance des structures cellulaires, etc.
En utilisant la DEB, les différences entre les espèces peuvent être réduites à des différences dans les valeurs d'énergie allouée aux différents paramètres. Cette théorie propose que le métabolisme dépend de la surface ou du volume de l’organisme plutôt que de sa masse, contrairement à la MTE, et trouve donc une relation entre métabolisme et volume avec un exposant 2/3 au lieu de ¾. Tout comme la MTE, la DEB cherche une organisation commune à tous les êtres vivants, et de nombreux contre-exemples viennent réfuter cette théorie.
La théorie métabolique soutient que les organismes partagent beaucoup de caractéristiques communes, notamment concernant leur métabolisme. Un exemple est le coefficient ¼ que l'on retrouve dans de nombreux taux biologiques (la croissance embryonnaire ou la vitesse de circulation du sang sont proportionnelles à M1/4, les diamètres des aortes des mammifères sont proportionnels à M3/4)[15]. Elle sous-entend donc qu'il existe des principes universels qui gouvernent l'évolution, et qu'ils sont étroitement liés aux contraintes énergétiques des organismes[15].
On retrouve là une hypothèse nommée « symmorphisme »[16], développée par Ewald R. Weibel (en) et Richard Taylor en 1981. D'après ces derniers, les systèmes biologiques tendent à adopter la structure la plus économique pour une fonction attribuée : par exemple les réseaux de capillaires, les muscles, les mitochondries, etc. sont bien « adaptées » au transfert de l'oxygène. Les structures n'auraient pas des capacités qui dépasseraient les besoins de l'organisme.
C'est principalement cet aspect qui est à l'origine du rejet de la théorie par certains auteurs, car elle questionne le caractère prévisible ou non de l'évolution.
Allen et Gillooly, cofondateurs de la théorie, reconnaissent cependant que la théorie a ses limites, et qu'en aucun cas elle n'a l'ambition d'expliquer tous les phénomènes biologiques. À l'avenir, les théoriciens espèrent pouvoir établir un lien plus évident entre l'écologie et l'évolution, via la théorie métabolique[15].
Notes et références
- Brown, J. H., Gillooly, J. F., Allen, A. P., Savage, V. M., & G. B. West (2004). "Toward a metabolic theory of ecology". The Ecological Society of America
- Geoffrey B. West, James H. Brown, and Brian J. Enquist, A General Model for the Origin of Allometric Scaling Laws in Biology, Science 4 April 1997: 276 (5309), 122-126. [DOI:10.1126/science.276.5309.122]
- O'Connor, M. P., Kemp, S. J., Agosta, S. J., Hansen, F., Sieg, A. E., Wallace, B. P., McNair, J. N. and Dunham, A. E. (2007), Reconsidering the mechanistic basis of the metabolic theory of ecology, Oikos, 116: 1058–1072. doi: 10.1111/j.0030-1299.2007.15534.x
- Kimberley Louwrens, (2011), http://www.bio.vu.nl/thb/deb/essays/Louw2011.pdf (essay)
- Gillooly JF, Brown JH, West GB, Savage VM, Charnov EL (2001) Effects of size and temperature on metabolic rate. Vol 293 Science .
- Gillooly, J. F, E. L. Charnov, G. B. West, V. M. Savage, and J. H. Brown. (2001) Effects of size and temperature on developmental time. Letters to Nature.
- Pauly, D. (1980) On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. Journal du Conseil international pour l'Exploration de la Mer(2): 175-192.
- Gillooly, J.F., Allen, A.P., West, G.B., & Brown, J.H. (2005). "The rate of DNA evolution: Effects of body size and temperature on the molecular clock". Proc Natl Acad Sci U S A
- Allen A.P., Brown J.H. & Gillooly J.F. (2002). "Global biodiversity, biochemical kinetics, and the energetic-equivalence rule". Vol 297 Science.
- Price CA, Weitz JS, Savage VM, Stegen J, Clarke A (2012) Testing the metabolic theory of ecology. Ecology Letters,
- Humphries, M. M., McCann, K. S. (2013), Metabolic ecology. Journal of Animal Ecology. doi: 10.1111/1365-2656.12124
- Bradford A. Hawkins, Fabio S. Albuquerque, Miguel B. Araújo, Jan Beck, Luis Mauricio Bini, Francisco J. Cabrero-Sañudo, Isabel Castro-Parga, José Alexandre Felizola Diniz-Filho, Dolores Ferrer-Castán, Richard Field, José F. Gómez, Joaquín Hortal, Jeremy T. Kerr, Ian J. Kitching, Jorge L. León-Cortés, Jorge M. Lobo, Daniel Montoya, Juan Carlos Moreno, Miguel Á. Olalla-Tárraga, Juli G. Pausas, Hong Qian, Carsten Rahbek, Miguel Á. Rodríguez, Nathan J. Sanders and Paul Williams, A Global Evaluation of Metabolic Theory as an Explanation for Terrestrial Species Richness Gradients, Ecology Vol. 88, No. 8 (Aug., 2007), pp. 1877-1888 Published by: Ecological Society of America
- Englund G, Ohlund G, Hein CL, Diehl S., (2011).Temperature dependence of the functional response, Ecology letters, doi: 10.1111/j.1461-0248.2011.01661.x.
- Knies JL, Kingsolver JG. (2010). Erroneous Arrhenius: modified arrhenius model best explains the temperature dependence of ectotherm fitness, The American Naturalist, doi: 10.1086/653662.
- 10. Allen, A. P. and Gillooly, J. F. (2007), The mechanistic basis of the metabolic theory of ecology. Oikos, 116: 1073–1077. doi: 10.1111/j.0030-1299.2007.16079.x
- (en) « Symmorphosis », sur encyclopedia.com (consulté le ).
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Metabolic theory of ecology » (voir la liste des auteurs).