Gènes homéotiques chez les animaux
La génétique évolutive révèle une grande plasticité des mécanismes développementaux. Parmi ces mécanismes, nous retrouvons les gènes homéotiques. Il s'agit de gènes qui interviennent dans le processus de développement pendant la phase embryonnaire, ils permettent la mise en place des structures dans l'organisme grâce à une activation spatio-temporelle très précise.
Découverte
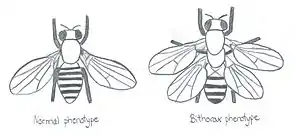
Pour bien comprendre la fonctionnement des gènes homéotiques, il faut remonter à la découverte des gènes homéotiques et ce, grâce aux observations des mutations dites homéotiques de certains gènes spécifiques, réalisées par le biologiste anglais William Bateson et accessibles dans son ouvrage publié en 1894[1].
La découverte de ces gènes homéotiques a été faite grâce aux observations sur des mutations dites homéotiques avec notamment l'exemple de mouches mutantes qui portent des pattes en lieu et place des antennes ou bien encore, la présence d'une seconde paire d'ailes au niveau d'un segment du thorax. Bateson baptise alors ce phénomène Homeosis[2].
Ce n'est qu'une centaine d'années plus tard que l'explication nous est donnée, par les prix Nobel de médecine 1995 qui sont l'Américain Edward B. Lewis, à l'Allemande Christiane Nüesslein-Volhard, ainsi qu'à l'Américain d'origine suisse Eric F. Wieschaus qui s'étaient alors intéressés alors à la relation entre gène homéotique et embryogenèse[2].
Il s'agit alors d'une mutation du gène qui va coder une protéine modifiée, qui ne pourra pas se lier à la séquence spécifique de l'ADN et ainsi modifier le développement spatio-temporel de la structure-cible. Dans le cas de l'homéose, il s'agit d'un développement spatial inadéquat expliqué par un mauvais positionnement de l'organe[2].
Généralités
Les gènes homéotiques influent sur le plan d'organisation d'un être vivant selon les axes de polarités qui sont l'axe antéro-postérieur (bouche-anus), l'axe dorso-ventral et l'axe bilatéral (gauche-droite). Il s'agit de gènes qui codent des protéines capables de se lier à des séquences spécifiques de l'ADN et ainsi influencer le développement de l'organisme au cours de l’embryogenèse (développement de l'individu dans l'embryon). Ils agissent donc comme des architectes contrôlant la structuration spatiale de l'organisme.
Les gènes homéotiques chez les vertébrés sont appelés gènes Hox. Les gènes Hox ont été identifiés à travers presque tout le règne animal, que soit chez l'homme, la souris, le serpent ou bien le poisson. Tous ces vertébrés possèdent des structures communes comme un système nerveux généralisé, avec le cerveau au niveau de la tête, et ensuite suivi du tronc cérébral et de la moelle épinière. C'est après que les gènes homéotiques entrent en jeu et permettent une individualisation des structures selon l'espèce. Cela est ancré dans le génome respectif de chaque espèce : la structure spatio-temporelle grâce aux gènes homéotiques est donc reproductible et héritée car elle définit au terme de l'embryogenèse les caractéristiques de l'espèce animale.
Il existe aussi des gènes Hox chez les invertébrés, et notamment chez les arthropodes comme la drosophile, qui est une espèce-modèle dans le milieu de la biologie et est très souvent utilisée comme modèle de base pour la génétique.
Fonctionnement
Les gènes homéotiques sont donc capables grâce à des protéines synthétisées de contrôler l'expression d'autres gènes. Ils agissent comme facteur de transcription car ils contrôlent par stimulation ou inhibition la transcription de gènes, c'est-à-dire le passage de l'ADN à l'ARN qui seront ensuite traduits en protéines. Ces gènes dits cibles assurent alors des fonctions essentielles à la bonne mise en place des structures[1].
Chez l'homme ou la souris, il existe 39 gènes homéotiques qui contribuent à l'établissement d'un axe antéro-postérieur bien généralisé. En effet, plusieurs études ont démontré que l'inactivation d'un gène homéotique contribue à un changement de l'axe antéro-postérieur. C'est notamment le cas chez la souris qui, dû à l'inactivation d'un gène Hox, a conduit à un changement de la morphologie de la deuxième vertèbre cervicale appelé axis en une vertèbre possédant la morphologie de l'atlas (première vertèbre cervicale qui porte le crâne). Cet exemple démontre donc l'importance des gènes Hox dans le développement de l'axe antéro-postérieur. Comme expliqué précédemment, si le gène homéotique n'est pas activé, alors il ne peut synthétiser des protéines permettant de contrôler l'expression d'autres gènes servant dans le développement[3].
Colinéarité
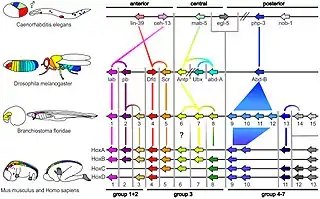
Il existe une relation qui lie l'ordre physique des gènes homéotiques au sein des complexes chromosomiques qui sont au nombre de 4 chez la souris, la localisation des territoires qui expriment les gènes Hox et le moment de leur expression, c'est ce qu'on appelle la colinéarité. Ainsi, les gènes occupant les premières positions au sein de leur complexe respectif seront les premiers à être exprimés au cours du développement embryonnaire et ce, au niveau les plus antérieurs. Ceux qui suivent mettent plus de temps avant d'être activés et donc avant d'être exprimés. Et ce mécanisme se poursuit jusqu'au dernier gène du complexe homéotique.
La structuration spatiale de l'organisme repose donc sur la colinéarité, qui détermine la programmation spatio-temporelle de l'expression des gènes Hox. Des expériences ont montré que si l'on perturbait la colinéarité, on affecte les structures en développement[4]. En reprenant l'exemple de la souris et de la morphologie de la seconde vertèbre cervicale en atlas au lieu de celle initiale axis, la colinéarité explique ce changement morphologique. En effet, alors que l'atlas n'exprime jamais le gène homéotique Hoxb4, l'axis, quant à lui, est censé l'exprimer. S'il n'est pas exprimé car inactivé, alors l'axis adopte la morphologie de l'atlas qui le précédait dans le développement. Pour résumer, l'absence du gène Hoxb4 empêche le développement de l'axis, qui prend alors la forme de son prédécesseur dans l'ordre spatial: la seconde vertèbre cervicale appelée axis devient comme la première vertèbre cervicale atlas. Ce mécanisme de colinéarité s'applique aussi bien chez les arthropodes où l'emplacement de chaque gène va déterminer son expression spatio-temporelle[3].
Détermination de l'expression des gènes Hox
Il existe des régulateurs de transcriptions des gènes Hox qui permettent d'établir les capacités de chacun de ces gènes homéotiques, d'expression spatio-temporelle. Il existe certaines molécules telle que l'acide rétinoïque qui dérive de la vitamine A ou certains autres facteurs de croissance, qui sont diffusés et se distribuent selon la position de certains organes en développement tels que le cerveau. Ces facteurs de croissance affectent alors les régulateurs de transcription qui peuvent alors activer les gènes Hox, et ce, à partir d'une certaine dose. Une localisation de l'axe antéro-postérieur de l'embryon est donc associée à une certaine concentration du facteur de croissance concernée[3].
Gènes Hox et organogenèse
Les gènes Hox qui sont impliqués dans l’embryogenèse sont par conséquent aussi impliqués dans l'organogenèse, c'est-à-dire la formation des organes. Ce sont les principaux architectes de la morphogenèse des systèmes respiratoire, digestif et urogénital. Chez les mammifères, et plus exactement l'espèce humaine, les gènes Hox des groupes 1 à 6 sont exprimés lors du développement de l'appareil respiratoire. Encore une fois, ces affirmations ont été démontrées grâce aux mutations exercées sur les gènes Hox en question. Ici, il s'agit donc des gènes Hoxa3 et Hoxa5 qui permettent la formation du larynx, et plus particulièrement l'utilisation du gène HoXa5 pour la formation de la trachée et du poumon. Tous les individus déficients pour le gène Hoxa5 présentent en effet une malformation des voies respiratoires[5].
Au niveau du développement digestif, l'exemple le plus probant est celui du tube digestif, il s'agit ici des gènes Hoxc4, Hoxa5, Hoxd12, Hoxa13 et Hoxd13 qui sont exprimés dans le mésoderme viscéral. Hoxc4 Est principalement lié au développement de la couche musculaire de l’œsophage, alors que les autres sont présents dans le tube digestif : Hoxd12 et Hoxd13 sont associés aux couches musculaires du rectum[5].
Enfin, au niveau du développement urogénital, c'est-à-dire ce qui est relatif à l'appareil génital et l'appareil urinaire, les gènes des groupes 10, 12 et 13 sont impliqués. Les mutations de Hoxa10, Hoxa11, Hoxd11, Hoxa13 et Hoxd13 entraînent une stérilité chez l'individu qui ne peut plus se reproduire. Notamment Hoxa10 et Hoxa11 entraînent le développement du canal déférent chez le mâle. Chez la femelle, le gène Hoxa10 participe à la formation de l'utérus[5].
Néanmoins, certaines particularités ne sont pas encore expliquées, comme le fait que ces développements ne sont pas associés à la colinéarité[5].
Post-natalité
Les gènes homéotiques sont souvent associés à l'embryogenèse, mais ils sont utilisés aussi après la naissance de l'individu. Leur impact post-natal n'est pas aussi conséquent que pendant la phase embryonnaire néanmoins certains gènes Hox sont utilisés par certains organes avec une utilisation spécifique. Ainsi, les gènes homéotiques sont impliqués notamment dans les hémopathies malignes. Encore une fois l'absence ou l'inactivité de certains gènes Hox permettent de confirmer l'hypothèse d'un impact post-natalité. Pour exemple, la triple mutation des gènes Hoxa9, Hoxb9 et Hoxd9 provoque une hypoplasie de la glande mammaire, caractérisée par la non-production de lait chez les femelles qui sont alors incapables de nourrir leurs petits[5].
Conclusion
Les gènes homéotiques participent donc principalement au bon développement des différentes structures de l'individu lors de l'embryogenèse, mais pas seulement, ils sont aussi essentiels à la bonne formation des organes essentiels à sa survie. Enfin, malgré leur prépondérance dans le développement embryonnaire, c'est-à-dire prénatal, les expériences démontrent aussi qu'ils influent sur le système des animaux même après leur naissance.
Notes et références
- Walter J. Gehring, La drosophile aux yeux rouges: gènes et développement,
- Jean Deutsch; Claudie Lamour-Isnard; Jean-Antoine Lepesant, « Le Prix Nobel 95 attribué à Ed Lewis, Christiane NüessleinVolhard et Eric Wieschaus: la reconnaissance de la génétique du développement », Medecine/Sciences,
- René Rezsohazy, « Gènes du développement et structuration de l’organisme », Revue des Questions scientifiques, , p. 99,110
- Jean Deustch, « Homéogènes », Médecines/sciences, , p. 401-403
- Josée Aubin; Lucie Jeannotte, « Implication des gènes Hox dans les processus d’organogenèse chez les mammifères », Médecine/Sciences,