Induction cellulaire chez les amphibiens au cours de la gastrulation
Une induction cellulaire est un processus par lequel un groupe de cellules engage un autre groupe de cellules adjacent dans une nouvelle voie de différenciation. Ce système intervient durant toutes les étapes de la vie d’un organisme : développement embryonnaire, post-embryonnaire et pendant la vie effective. Lors de la reproduction sexuée, après fécondation, il y a développement de l’organisme. L’ontogénèse est l’ensemble des éléments permettant de passer d’une cellule œuf à un organisme adulte. Elle comprend le développement embryonnaire :
- Segmentation
- Gastrulation
- Neurulation
- Organogénèse
Et post-embryonnaire :
- Croissance
- Métamorphose
La gastrula vient du mot grec « estomac » et permet la mise en place des trois feuillets embryonnaires primordiaux définissant les triblastiques, groupe des amphibiens ;
- Ectoderme externe qui donnera l’épiderme et le système nerveux
- Mesoderme intermédiaire qui donnera les muscles, les vaisseaux sanguins et le squelette
- Endoderme interne qui donnera les glandes annexes et le tube digestif.
Les amphibiens appartiennent au groupe des métazoaires et sont utilisés comme modèle scientifique car leur développement est rapide et visible à l’échelle macroscopique. L'induction cellulaire aura donc un rôle clef au cours de la gastrulation, car elle met en contact les tissus inducteurs avec les tissus voués à se différencier. Les signaux seront de diverse nature chimique et agiront sur diverses composantes cellulaires qui leur sont spécifique (protéines, ADN, récepteurs transmembranaires). L'objectif final étant la mise en place du plan d'organisation du xénope, avec des tissus différentiés et fonctionnels.
Mise en place
La reproduction des amphibiens donne un œuf hétérolécithe. Le spermatozoïde va féconder l’ovocyte au niveau du pôle animal, et ainsi déclencher une cascade de réactions cytoplasmiques. Le cortex bascule, entrainant la répartition différentielle des déterminants cytoplasmiques maternelles : accumulation de β-caténine et Vg1 dans le centre de Nieuwkoop. De plus le cortex entraîne avec lui un pigment, laissant une zone claire dépigmentée, appelée le croissant gris. Ce dernier est un centre organisateur car il est à l’origine des lèvres blastoporales qui mettent en contact des groupes de cellules sécrétant les messages chimiques permettant l’induction et donc la mise en place des feuillets. L’induction cellulaire pendant la gastrulation est une alternance entre l’expression de certains gènes à des périodes précises corrélées avec des phases de migration cellulaire. Il y a chez les amphibiens des gènes sélecteurs homéotiques, qui sélectionnent les gènes actifs dans chaque segment et s’expriment dès la gastrulation. Ils suivent la loi de colinéarité (3’ → 5’), et ils sont localisés dans le même ordre que les futures parties du corps de l’amphibien (tête → queue).
Induction et structuration du mésoderme
Induction verticale des blastomères
Elle est mise en évidence par les expériences de Nieuwkoop. Au stade blastula (32 cellules) on peut montrer qu’il existe des signaux inducteurs issus des blastomères végétatifs qui vont agir sur des blastomères des cellules animales, afin de changer leur devenir. Les cellules de la zone marginale vont acquérir la capacité à se différencier en cellules mésodermiques. On dit qu’elles sont déterminées à former du mésoderme. Il existe donc un processus d’induction verticale, selon la position de l’axe pôle animal-pôle végétatif vertical.
Induction horizontale
Elle est mise en évidence par les expériences de Dale et Slack grâce à la recombinaison des bastomères. La position dorso-ventrale des blastomères végétatifs apparaît comme étant fondamentale dans la détermination des cellules de la zone marginale à former un type spécifique de mésoderme. Il existe une induction dorso-ventrale c'est-à-dire que l’induction issue des blastomères végétatifs détermine une régionalisation dorso-ventrale du mésoderme.
Les blastomères dorso-végétatifs forment un centre inducteur : le centre de Nieuwkoop
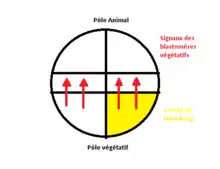
Il est mis en évidence grâce aux expériences de Gimlich et Gerhart. Les blastomères végétatifs qui sont capables d’induire un axe embryonnaire sans participer directement à la formation de cet axe, forment le centre de Nieuwkoop. Ces blastomères végétatifs vont hériter de déterminants maternels à potentialité dorsale.
Ainsi l'induction cellulaire se fait selon l'axe pôle animal-pôle végétatif de l'embryon et participe à la mise en place du mésoderme ainsi que de la position dorso-ventrale des tissus issus de la gastrulation.
Interactions moléculaires
Une induction est un processus par lequel un groupe de cellules engage un autre groupe de cellules adjacent dans une nouvelle voie de différenciation. Ici les cellules adjacentes sont les blastomères ou d’autres cellules devenues adjacentes lors des mouvements morphogénétiques qui les ont rapprochées lors de la gastrulation.
Facteurs de transcription
Un facteur de transcription est une molécule capable de se fixer sur la région promotrice d’un gène afin de réguler son expression. On trouve un certain nombre de facteurs de transcription ayant un rôle fondamental dans l'induction au cours de la gastrulation chez les amphibiens:
- La β-caténine, d'origine maternelle, s’accumule dans les blastomères dorso-végétatifs lors de la rotation corticale. Au cours de la segmentation elle va être adressée vers les noyaux des blastomères dorso-végétatifs et sa localisation sera toujours restreinte au centre de Nieuwkoop, dont elle peut mimer l'action. En se fixant sur le promoteur du gène Siamois, elle active sa transcription et induit un axe embryonnaire surnuméraire.
- VegT, qui est plus précoce, localisé dans le cytoplasme cortical de l'hémisphère végétatif. Il joue un rôle fondamental dans l’induction verticale du mésoderme, et participe également à l'autodétermination de l'endoderme.
- Goosecoïd, facteur de transcription d'origine zygotique et exprimé par le centre de Spemann, donnant du mésoderme dorsal à l'origine de la chorde.
- Brachyury, également d'origine zygotique, exprimé dans tout le mésoderme, induit la formation des muscles. C'est un marqueur pan-mésodermique.
TGF : Transforming Growth Factor
Ce sont des petits peptides qui ont tous la même modalité d’action. Ils induisent des cascades de réaction au sein de la cellule afin d’agir sur l’ADN. On trouve parmi eux :
- Vg1 : c’est un déterminant maternel qui fait partie des premiers signaux inducteurs. Il est situé dans la zone corticale grâce à la rotation corticale, et induit le mésoderme dorsal.
- Activines et Nodal : ce sont des TGFβ sécrétés d'origine zygotique, exprimés selon un gradient dorso-ventral. Ils induisent le mésoderme par effet dose-dépendant.
- Bone Morphogenetic Protein (BMP4) : il s'agit d'un TGFβ à effet ventralisant du mésoderme, exprimé par le mésoderme ventral. Il induit également l'épiderme.
Les FGF : Fibroblast Growth Factor
Ce sont des facteurs de croissance toujours présents chez l’adulte qui assurent la prolifération des cellules. Ils seraient d'origine maternelle. Ils sont sécrétés au niveau du centre inducteur ventral (endoderme ventro-latéral). Chez l’embryon, leurs principaux rôles sont :
- Le changement de forme des cellules.
- La modification de l’adhésivité des cellules.
- Rôle amplificateur des réactions.
- Induction primaire du mésoderme ventral, postérieur et latéral.
Ce sont des signaux modulateurs, qui activent une transduction du signal vio un récepteur à activité tyrosine-kinase. On trouve parmi eux : Noggin, Chordin, Follistatin et Wnt.
Mode d’action des facteurs inducteurs


Gradients de concentration dorso-ventraux
Les molécules présentées ci-dessus ont des concentrations différentes qui vont permettre la formation d’organes via l’intervention de molécules chimiques ou de facteurs de transcription :
- À forte concentration en région ventrale, l’activine induit le mésoderme ventro-latéral, et à forte concentration en région dorsale induit le mésoderme dorsal. À une concentration moyenne elle active brachyury.
- Vg1 est peu concentré en région ventrale ce qui induit le mésoderme ventral. À l’inverse en région dorsale il est fortement concentré ce qui engendre la formation du mésoderme dorsal.
- BMP4 a son gradient opposé à celui de l’activine. Ceci permet la régulation des signaux inducteurs, et la mise en place précise des feuillets.
L’induction est donc contrôlée par des gradients de concentration dorso-ventraux, et la concentration même des molécules inductrices permet la régulation et donc la mise en place des feuillets, primordiale dans le développement d'un individu possédant un plan d’organisation qui lui est propre ainsi que des organes fonctionnels. Il s’agit d’un contrôle spatial indiquant une information de position.
Cascades de réactions déclenchées
Les molécules agissent également dans le temps, permettant l’acquisition de compétences. L’induction des cellules n’est possible que dans un temps limité. On parle de cascade de réactions permettant la mise en place des feuillets.
- L’induction verticale du mésoderme: les signaux chimiques sont envoyés par les blastomères du pôle végétatif vers la zone marginale. Ils sont envoyés par des déterminants maternelles tels que Veg T et sont amplifiés par les FGF qui permettent l’expression des gènes cibles.
- Régionalisation du centre de Nieuwkoop: l’activine, Vg1, la β-caténine agissant sur Nodal et permettant l’expression de Chordin sont localisés dans le centre de Nieuwkoop. Ceci engendre la prolifération des cellules, leur changement de forme ainsi que leur cohésion.
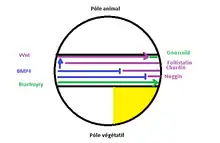 Cascade de réactions déclenchées
Cascade de réactions déclenchées - Une troisième vague dans le centre de Spemann.
- Brachyury s’exprime dans toutes le mésoderme.
- BMP4 s’exprime en région ventrale et permet un contrôle négatif sur Noggin et Chordin afin qu’ils s’expriment en région dorsale. Il active également Wnt.
- Wnt agit sur goosecoïd permettant de restreindre son expression en région dorsale.
Notes et références
- Bases cellulaires et moléculaires du développement, Christophe Chanoine, éditions Ellipses
- Biologie du développement. Les grands principes, L.WOLPERT, éd.DUNOD