Gradient latitudinal de biodiversité
Le Gradient latitudinal de biodiversité, GLB (de l'anglais latitudinal diversity gradient, LDG ou latitudinal biodiversity gradient, LBG) est un paramètre environnemental censé rendre compte de l'augmentation de la richesse en espèces, ou biodiversité, des pôles aux tropiques laquelle se produit pour une grande variété d'organismes terrestres et marins [1]. Le GLB, l'un des modèles les plus largement reconnus en écologie[1], a été observé à des degrés divers dans l’histoire biologique de la Terre[2]. Une tendance parallèle a été observée avec l'altitude (gradient altimétrique de biodiversité )[3], bien que cela soit moins bien étudié[4].
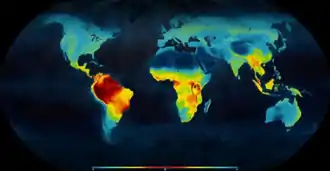
(Mannion 2014)
Expliquer le gradient latitudinal de biodiversité est considéré comme l’un des grands défis contemporains de la biogéographie et de la macroécologie (Willig et al. 2003, Pimm et Brown 2004, Cardillo et al. 2005)[5]. La question « Qu'est-ce qui détermine les modèles de diversité des espèces ? » figurait parmi les 25 principaux thèmes de recherche pour l'avenir identifiés dans le numéro du 125e anniversaire de Science en juillet 2005. Il y a pas de consensus parmi les écologistes sur les mécanismes sous-jacents au modèle, et de nombreuses hypothèses ont été proposées et débattues. Une étude de 2003 a noté que parmi les nombreuses énigmes associées au GLB, la relation causale entre les taux d'évolution moléculaire et la spéciation reste à démontrer[6].
Comprendre la répartition mondiale de la biodiversité est l'un des objectifs les plus importants pour les écologistes et les biogéographes. Au-delà des objectifs purement scientifiques et de la satisfaction de la curiosité, cette compréhension est essentielle pour des questions pratiques d'importance majeure pour l'humanité, telles que la propagation des espèces envahissantes, le contrôle des maladies et de leurs vecteurs, et les effets probables du changement climatique mondial sur le maintien de la biodiversité (Gaston 2000). Les zones tropicales jouent un rôle de premier plan dans la compréhension de la répartition de la biodiversité, car le taux de dégradation de leur habitat et de leur perte de biodiversité sont exceptionnellement élevés[7].
Le modèle dans le passé
Si le GLB est un modèle notable chez les organismes actuels, décrit qualitativement et quantitativement, étudié à divers niveaux taxonomiques, à travers différentes périodes et dans de nombreuses régions géographiques (Crame 2001), il a également pu être observé à des degrés divers dans les âges anciens, probablement en raison de différences de climat au cours des différentes phases de l'histoire de la Terre. Certaines études indiquent qu'il y avait un fort GLB, particulièrement dans les taxons marins ; par contre d'autres études indiquent que le GLB avait peu d'effet sur la distribution des animaux des taxons terrestres[2].
Hypothèses pour étayer le modèle
Bien que les nombreuses hypothèses visant à explorer le GLB soient étroitement liées et interdépendantes, la plupart d'entre elles peuvent être regroupées en trois hypothèses générales. Certaines de ces hypothèses peuvent elles-mêmes se subdiviser en hypothèses de second niveau :
- Les hypothèses liées aux espaces géographiques :
- Hypothèse de l'effet de domaine intermédiaire ;
- Hypothèse de l'aire géographique ;
- Hypothèse espèce-énergie ;
- Hypothèse de la dureté du climat ;
- Hypothèse de stabilité climatique.
- Les hypothèses liées à l'évolution :
- Hypothèse de la perturbation historique ;
- Hypothèse du taux d'évolution ;
- Hypothèse du temps évolutif effectif.
- Les hypothèses biotiques c'est-à-dire liées à la biologie des espèces.
Hypothèses spatiales / superficielles
Cinq hypothèses majeures se basent uniquement sur les caractéristiques spatiales et les propriétés des aires de répartition géographiques des tropiques :
Hypothèse de l'effet de domaine intermédiaire
Cette hypothèse repose sur les modèles informatiques de Cowell et Hurt (1994) ainsi que de Willing et Lyons (1998) qui semblent montrer que, si les aires de répartition des espèces étaient mélangées au hasard à l'intérieur d'un domaine géographique délimité exempt de gradients environnementaux, les aires de répartition des espèces auraient tendance à se chevaucher davantage vers le centre du domaine que vers ses limites, créant un pic de richesse spécifique « à mi-domaine ». Ce constat qui a été qualifié d'« effet de domaine intermédiaire » (Mid-Domain Effect, MDE) est controversé à la fois dans son concept que dans son application réelle[5] - [8], certains auteurs rapportent en effet qu'il y a peu de correspondance entre les modèles de diversité latitudinale prévus et observés (Bokma and Monkkonen 2001, Currie and Kerr, 2007, Diniz-Filho et al. 2002, Hawkins and Diniz-Filho 2002, Kerr et al. 2006).
Hypothèse de l'aire géographique
Terborgh (1973) affirme que les zones tropicales constituant le plus grand biome, ces zones peuvent abriter davantage d'espèces ; celles-ci ayant des taux d'extinction plus faibles, (Rosenzweig 2003), elles seraient donc plus susceptibles de subir une spéciation allopatrique, ce qui augmenterait leur taux de spéciation (Rosenzweig 2003). La combinaison entre taux d'extinction plus faibles et taux élevés de spéciation conduirait à des niveaux élevés de richesse en espèces dans les zones tropicales.
Une critique de cette hypothèse est que même si les tropiques sont le plus étendu des biomes, les biomes successifs au nord des tropiques ont tous à peu près la même superficie. Ainsi, si l'hypothèse de l'aire géographique est correcte, ces régions devraient toutes avoir à peu près la même richesse en espèces, ce qui n'est pas le cas comme l'indique le fait que les régions polaires contiennent moins d'espèces que les régions tempérées (Gaston et Blackburn 2000). Pour expliquer cela, Rosenzweig (1992) a suggéré que si les espèces à répartition partiellement tropicale étaient exclues, le gradient de richesse au nord des tropiques disparaîtrait. Blackburn et Gaston 1997 ont testé l'effet de l'élimination des espèces tropicales sur les modèles latitudinaux de la richesse en espèces aviaires dans le Nouveau Monde et ont constaté qu'il existe effectivement une relation entre la superficie terrestre et la richesse en espèces d'un biome une fois que les espèces à prédominance tropicale sont exclues.
Peut-être un défaut plus grave dans cette hypothèse est l'argument avancé par certains biogéographes qui suggèrent que les tropiques terrestres ne sont pas, en fait, le plus grand biome, et donc cette hypothèse ne serait pas une explication valable au gradient latitudinale de diversité des espèces (Rohde 1997, Hawkins et Porter 2001). En tout état de cause, il serait difficile de défendre les tropiques en tant que « biome » plutôt que les régions géographiquement diverses et disjointes qu'elles englobent réellement.
Il a été démontré que l'effet de la superficie sur les modèles de biodiversité dépend de l'échelle, cette dernière ayant une conséquence plus forte sur les espèces à petites aires géographiques par rapport aux espèces à grandes aires de répartition lesquelles sont davantage affectées par d'autres facteurs tels que le "domaine intermédiaire" et / ou la température[5].
Hypothèse espèce-énergie
L'hypothèse espèce-énergie suggère que la quantité d'énergie disponible est un facteur limitant de la richesse du système. Ainsi, une énergie solaire accrue (avec une abondance d'eau) aux basses latitudes entraîne une augmentation de la productivité primaire nette (issue de la photosynthèse). Cette hypothèse propose que plus la productivité primaire nette est élevée, plus il y aura d'individus et plus il y aura d'espèces dans une zone. En d'autres termes, cette hypothèse suggère que les taux d'extinction sont réduits vers l'équateur en raison de populations plus nombreuses soutenues par une plus grande quantité d'énergie disponible dans les zones tropicales ; dans ces zones les taux d'extinction plus faibles conduiraient à l'émergence d'un plus grand nombre d'espèces.
Hypothèse de la dureté du climat
Cette hypothèse liée au climat stipule que le GLB peut exister simplement parce que peu d'espèces peuvent physiologiquement tolérer des conditions à des latitudes plus élevées qu'aux basses latitudes, car les premières sont plus froides et plus sèches que les secondes. Currie et al. (2004) ont critiqué cette hypothèse en affirmant que, s'il est clair que la tolérance climatique peut limiter la répartition des espèces, il apparaît que certaines espèces sont souvent absentes des zones dont elles pourraient tolérer le climat[9].
Hypothèse de stabilité climatique
À l'instar de l'hypothèse de la dureté du climat, la stabilité du climat serait à l'origine du gradient de biodiversité. Le raisonnement qui conduit à cette hypothèse consiste à supposer que si un environnement fluctuant est susceptible d'augmenter le taux d'extinction ou d'empêcher la spécialisation, un environnement constant peut permettre aux espèces de se spécialiser sur des ressources alimentaires prévisibles, leur permettant d'avoir des niches plus étroites et ainsi faciliter leur spéciation. Le fait que les régions tempérées ont des conditions saisonnières plus variables sur les échelles de temps géologiques suggère que les régions tempérées devraient donc avoir moins de diversité d'espèces que les tropiques.
Les critiques de cette hypothèse soulignent le fait qu'il existe de nombreuses exceptions à l'hypothèse selon laquelle la stabilité du climat signifie une plus grande diversité des espèces. Par exemple, on sait que la faible diversité des espèces se produit souvent dans des environnements stables tels que le sommet des montagnes tropicales. De plus, de nombreux habitats présentant une grande diversité d'espèces tout en vivant sous des climats saisonniers, y compris de nombreuses régions tropicales lesquelles ont des précipitations très variables selon l'époque de l'année (Brown et Lomolino 1998).
Hypothèses historiques et évolutives
Trois hypothèses principales sont liées aux explications historiques et évolutives de l'augmentation de la diversité des espèces vers l'équateur :
L'hypothèse de la perturbation historique
L'hypothèse des perturbations historiques propose que la faible richesse en espèces des latitudes plus élevées est une conséquence d'une période de temps insuffisante pour que les espèces colonisent ou recolonisent des zones en raison de perturbations historiques telles que la glaciation (Brown et Lomolino 1998, Gaston et Blackburn 2000). Cette hypothèse suggère que la diversité dans les régions tempérées n'a pas encore atteint l'équilibre et que le nombre d'espèces dans les régions tempérées continuera d'augmenter jusqu'à saturation (Clarke et Crame 2003).
L'hypothèse du taux d'évolution
L'hypothèse du taux d'évolution soutient que des taux d'évolution plus élevés sous les tropiques ont entraîné des taux de spéciation plus élevés et donc une diversité accrue aux basses latitudes (Cardillo et al.2005, Weir & Schluter 2007, Rolland et al. 2014). Des taux d'évolution plus élevés sous les tropiques ont été attribués à des températures ambiantes plus élevées, des taux de mutation plus élevés, un temps de génération plus court et / ou des processus physiologiques plus rapides (Rohde 1992, Allen et al.2006) et une pression de sélection accrue d'autres espèces qui évoluent elles-mêmes[10]. Des taux plus rapides de microévolution dans les climats chauds (c.-à-d. basses latitudes et altitudes) ont été observés pour les plantes (Wright et al. 2006), les mammifères (Gillman et al. 2009) et les amphibiens (Wright et al. 2010). Si l'on suppose que des taux de microévolution plus rapides entraînent des taux de spéciation plus rapides, ces résultats suggèrent que des taux d’évolution plus rapides dans les climats chauds ont presque certainement une forte influence sur le GLB. Des recherches supplémentaires doivent être menées pour déterminer si les taux de spéciation sont effectivement plus élevés sous les tropiques. Comprendre si le taux d'extinction varie avec la latitude sera également important pour savoir si cette hypothèse est ou non étayée (Rolland et al. 2014).
L'hypothèse du temps évolutif effectif
L'hypothèse du temps évolutif effectif suppose que la diversité est déterminée par le temps évolutif pendant lequel les écosystèmes ont existé dans des conditions relativement inchangées, et par la vitesse d'évolution directement déterminée par les effets de l'énergie environnementale (température) sur les taux de mutation, les temps de génération et la vitesse de sélection (Rohde 1992). Elle diffère de la plupart des autres hypothèses en ne postulant pas une limite supérieure de la richesse en espèces fixée par divers facteurs abiotiques et biotiques ; en d'autres termes il s'agit d'une hypothèse de non-équilibre supposant un espace de niche largement non saturé. Rohde admet que de nombreux autres facteurs peuvent également jouer un rôle dans la formation de gradients latitudinaux de richesse en espèces. L'hypothèse est cependant étayée par de nombreuses preuves, en particulier les études d'Allen et al. (2006) et Wright et al. (2006).
Hypothèses biotiques
Les hypothèses biotiques affirment que les interactions écologiques des espèces telles que la compétition, la prédation, le mutualisme et le parasitisme sont plus fortes sous les tropiques et ces interactions favorisent la coexistence des espèces et leur spécialisation, conduisant à une plus grande spéciation dans les tropiques. Ces hypothèses sont problématiques car elles ne peuvent pas être la cause immédiate du GLB car elles ne parviennent pas à expliquer pourquoi les interactions entre les espèces seraient plus fortes sous les tropiques.
Un exemple d'une telle hypothèse affirme que la plus grande intensité de la prédation et l'existence de prédateurs plus spécialisés dans les tropiques ont contribué à l'augmentation de la diversité dans ces zones (Pianka 1966). Cette intense prédation pourrait réduire l'importance de la concurrence (voir principe d'exclusion compétitive), permettre un plus grand chevauchement des niches et favoriser une plus grande richesse de proies. Alors que des expériences à grande échelle suggèrent que la prédation pourrait être plus intense sous les tropiques[11] - [12] cela ne peut pas être la cause ultime d'une grande diversité tropicale car cela ne permet pas d'expliquer ce qui donne lieu à la richesse des prédateurs sous les tropiques.
Plusieurs études n'ont pas réussi à observer des changements cohérents dans les interactions écologiques en fonction de la latitude (Lambers et al. 2002)[1]. Ces études suggèrent que l'intensité des interactions entre les espèces n'est pas corrélée avec le changement de la richesse des espèces selon la latitude.
Synthèse et conclusions
Il existe de nombreuses autres hypothèses liées au GLB, mais les hypothèses évoquées donnent un bon aperçu des enjeux débattus. Il est important de noter que bon nombre de ces hypothèses sont similaires et dépendent les unes des autres. Par exemple, les hypothèses évolutives dépendent étroitement des caractéristiques climatiques historiques des tropiques.
La généralité du gradient de diversité latitudinale
Une méta-analyse approfondie de près de 600 gradients latitudinaux provenant de la littérature publiée a testé la généralité du GLB à travers différents organismes, habitats et caractéristiques régionales[1]. Les résultats ont montré que le gradient latitudinal se produit en milieu marin, les écosystèmes terrestres et d’eau douce, dans les deux hémisphères. Le gradient est plus prononcé chez les taxons plus riches (c'est-à-dire les taxons ayant le plus d'espèces), les organismes plus grands, dans les écosystèmes marins et terrestres par rapport aux écosystèmes d'eau douce, et aux échelles régionale. La pente du gradient (la quantité de changement de la richesse en espèces avec la latitude) n'est pas influencée par la dispersion, le niveau trophique attaché à la physiologie animale (homéothermie ou ectothermie), l'hémisphère nord ou sud ou la plage latitudinale d'étude.
Les recherches n'ont pas pu directement mettre en défaut ou privilégier l'une des hypothèses ci-dessus, cependant, les résultats des études menées suggèrent qu'une combinaison énergie / climat et caractéristiques physiques des zones, contribuent probablement au gradient latitudinal des espèces. Les exceptions notables à cette tendance comprennent les insectes ichneumonidae, les oiseaux de rivage, les manchots et le zooplancton d'eau douce.
Robustesse des données
L'un des principaux postulas concernant les GLB et les modèles de richesse en espèces est que les données sous-jacentes (c'est-à-dire les listes d'espèces à des emplacements spécifiques) sont complètes. Cependant, dans la plupart des cas, ce postula n'est pas satisfait. Par exemple, les schémas de diversité des parasites sanguins des oiseaux suggèrent une plus grande diversité dans les régions tropicales, cependant, les données peuvent être faussées par un sous-échantillonnage dans des zones fauniques riches telles que l'Asie du Sud-Est et l'Amérique du Sud[13]. Pour les poissons marins, qui font partie des groupes taxonomiques les plus étudiés, les listes actuelles d'espèces sont extrêmement incomplètes pour la plupart des océans du monde. À une résolution spatiale de 3° (environ 350 km2), moins de 1,8% des océans du monde ont plus de 80% de leur faune piscicole actuellement décrite[14].
Conclusion
La question macroécologique fondamentale dont dépend la théorie du GLB est « Qu'est-ce qui cause les modèles de richesse en espèces ? ». La richesse en espèces dépend en fin de compte de tous les facteurs immédiats qui affectent les processus de spéciation, d'extinction, d'immigration et d'émigration. Alors que certains écologistes continuent de rechercher le mécanisme primaire ultime qui cause le gradient latitudinal de richesse biologique, de nombreux écologistes suggèrent plutôt que ce modèle écologique est susceptible d'être généré par synergie de plusieurs mécanismes (Gaston et Blackburn 2000, Willig et al.2003, Rahbek et al. 2007). Pour l'instant, le débat sur la cause du GLB se poursuivra jusqu'à ce qu'une étude révolutionnaire fournisse des preuves concluantes, ou qu'il y ait un consensus général sur le fait que de multiples facteurs contribuent à ce phénomène.
Notes et références
- Références
- (en) Helmut Hillebrand, « On the Generality of the Latitudinal Diversity Gradient », The American Naturalist, vol. 163, no 2, , p. 192–211 (ISSN 0003-0147, e-ISSN 1537-5323, PMID 14970922, DOI 10.1086/381004)
- (en) S.Sahney et M.J. Benton, « Recovery from the most profound mass extinction of all time », Proceedings of the Royal Society-B, vol. 275, no 1636, , p. 759–65 (ISSN 0962-8452, PMID 18198148, PMCID 2596898, DOI 10.1098/rspb.2007.1370)
- (en) Christy M. McCain, « Elevational Gradients in Diversity of Small Mammals », Ecology, vol. 86, no 2, , p. 366–372 (ISSN 0012-9658, DOI 10.1890/03-3147)
- (en) C. Rahbek, « The elevational gradient of species richness: a uniform pattern? », Ecography, vol. 18, no 2, , p. 200-205 (ISSN 0906-7590, e-ISSN 1600-0587, DOI 10.1111/j.1600-0587.1995.tb00341.x)
- (en) C. Mora & D.R. Robertson, « Causes of latitudinal gradients in species richness: a test with fishes of the Tropical Eastern Pacific », Ecology, vol. 86, no 7, , p. 1771–1792 (ISSN 0012-9658, DOI 10.1890/04-0883, lire en ligne, consulté le )
- (en) E. J. Dowle & al., « Molecular evolution and the latitudinal biodiversity gradient », Heredity, vol. 110, no 6, , p. 501–510 (ISSN 0018-067X, e-ISSN 1365-2540, PMID 23486082, PMCID 3656639, DOI 10.1038/hdy.2013.4)
- (en) D.Tittensor, « Global patterns and predictors of marine biodiversity across taxa », Nature, vol. 466, no 7310, , p. 1098–1101 (ISSN 0028-0836, e-ISSN 1476-468[à vérifier : ISSN invalide], PMID 20668450, DOI 10.1038/nature09329, lire en ligne, consulté le )
- Mora C, Sale, Kritzer et Ludsin, « Patterns and processes in reef fish diversity », Nature, vol. 421, no 6926, , p. 933–936 (PMID 12606998, DOI 10.1038/nature01393, Bibcode 2003Natur.421..933M, lire en ligne)
- D. J. Currie, G. G. Mittelbach, H. V. Cornell, D. M. Kaufman, J. T. Kerr et T. Oberdorff, « Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness », Ecology Letters, vol. 7, no 11, , p. 1121–1134 (DOI 10.1111/j.1461-0248.2004.00671.x)
- (en) Douglas W. Schemske, Gary G. Mittelbach, Howard V. Cornell, James M. Sobel et Kaustuv Roy, « Is There a Latitudinal Gradient in the Importance of Biotic Interactions? », Annual Review of Ecology, Evolution, and Systematics, vol. 40, no 1, , p. 245–269 (ISSN 1543-592X, DOI 10.1146/annurev.ecolsys.39.110707.173430)
- (en) Tomas Roslin et al., « REPORT Higher predation risk for insect prey at low latitudes and elevations (Risque de prédation plus élevé pour les proies des insectes aux basses latitudes et altitudes) », Science, vol. 356, no 6339, , p. 742–744 (ISSN 0036-8075, PMID 28522532, DOI 10.1126/science.aaj1631, Bibcode 2017Sci...356..742R, lire en ligne, consulté le )
- (en) A. L. Hargreaves et al., « Seed predation increases from the Arctic to the Equator and from high to low elevations (La prédation des semences augmente de l'Arctique à l'équateur et de haute à basse altitude) », Science Advances, vol. 5, no 2, , p. 4403 (ISSN 2375-2548, PMID 30801010, PMCID 6382403, DOI 10.1126/ sciadv.aau4403, Bibcode 2019SciA .... 5.4403 H)
- Nicholas Clark, Clegg, S. et Lima, M., « Une revue de la diversité mondiale chez les hémosporidies aviaires (Plasmodium et Haemoproteus: Haemosporida): nouvelles perspectives à partir de données moléculaires », International Journal for Parasitology, vol. 44, no 5, , p. 329–338 (PMID 24556563, DOI 10.1016 / j.ijpara.2014.01.004, hdl 10072/61114
 )
) - Mora C et Myers, « L'exhaustivité des inventaires taxonomiques pour décrire la diversité et la distribution mondiales des poissons marins », Actes de la Royal Society B, vol. 275, no 1631, , p. 149–155 (PMID 17999950, PMCID 2596190, DOI 10.1098 / rspb.2007.1315, http : //www.soc.hawaii.edu/mora/Publications/Mora%20019.pdf)
- Bibliographie
- A. P. Allen, J. F. Gillooly, V. M. Savage et J. H. Brown, « Kinetic effects of temperature on rates of genetic divergence and speciation », PNAS, vol. 103, no 24, , p. 9130–9135 (PMID 16754845, PMCID 1474011, DOI 10.1073/pnas.0603587103, Bibcode 2006PNAS..103.9130A)
- T. M. Blackburn et K. J. Gaston, « The relationship between geographic area and the latitudinal gradient in species richness in New World birds », Evolutionary Ecology, vol. 11, no 2, , p. 195–204 (DOI 10.1023/A:1018451916734)
- F. J. Bokma, M. Monkkonen et Mikko Mönkkönen, « Random processes and geographic species richness patterns: why so few species in the north? », Ecography, vol. 24, , p. 43–49 (ISSN 0906-7590, DOI 10.1034/j.1600-0587.2001.240106.x)
- Brown, J. H., and M. V. Lomolino. 1998. Biogeography. Sinauer Associates, Sunderland.
- M. Cardillo, C. D. L. Orme et I. P. F. Owens, « Testing for latitudinal bias in diversification rates: An example using New World birds », Ecology, vol. 86, no 9, , p. 2278–2287 (DOI 10.1890/05-0112)
- Clarke, A., and J. A. Crame. 2003. The importance of historical processes in global patterns of diversity. Pages 130-151 in T. M. Blackburn and K. J. Gaston, editors. Macroecology Concepts and Consequences. Blackwell Scientific, Oxford.
- R. K. Colwell et G. C. Hurtt, « Nonbiological gradients in species richness and a spurious Rapoport effect », American Naturalist, vol. 144, no 4, , p. 570–595 (DOI 10.1086/285695)
- R. K. Colwell et D. C. Lees, « The mid-domain effect: geometric constraints on the geography of species richness », Trends in Ecology & Evolution, vol. 15, no 2, , p. 70–76 (PMID 10652559, DOI 10.1016/s0169-5347(99)01767-x)
- R. K. Colwell, C. Rahbek et N. Gotelli, « The mid-domain effect and species richness patterns: what have we learned so far? », American Naturalist, vol. 163, no 3, , E1–E23 (PMID 15026983, DOI 10.1086/382056)
- R. K. Colwell, C. Rahbek et N. Gotelli, « The mid-domain effect: there's a baby in the bathwater », American Naturalist, vol. 166, no 5, , E149–E154 (DOI 10.1086/491689, lire en ligne)
- S. R. Connolly, « Process-based models of species distributions and the mid-domain effect », American Naturalist, vol. 166, no 1, , p. 1–11 (PMID 15937785, DOI 10.1086/430638, lire en ligne)
- J. A. Crame, « Taxonomic diversity gradients through geological time », Diversity and Distributions, vol. 7, no 4, , p. 175–189 (DOI 10.1046/j.1472-4642.2001.00106.x)
- D. J. Currie et J. T. Kerr, « Testing, as opposed to supporting, the Mid-domain Hypothesis: a response to Lees and Colwell (2007) », Ecology Letters, vol. 10, no 9, , E9–E10 (DOI 10.1111/j.1461-0248.2007.01074.x)
- D. J. Currie, G. G. Mittelbach, H. V. Cornell, D. M. Kaufman, J. T. Kerr et T. Oberdorff, « Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness », Ecology Letters, vol. 7, no 12, , p. 1121–1134 (DOI 10.1111/j.1461-0248.2004.00671.x)
- J. A. Diniz-Filho, T. F. L. V. B. Rangel, Marcia Christianne De Souza et Thiago F. L. V. B. Rangel, « Null models and spatial patterns of species richness in South American birds of prey », Ecology Letters, vol. 5, , p. 47–55 (DOI 10.1046/j.1461-0248.2002.00289.x)
- R. R. Dunn, C. M. McCain et N. Sanders, « When does diversity fit null model predictions? Scale and range size mediate the mid-domain effect. », Global Ecology and Biogeography, vol. 16, no 3, , p. 305–312 (DOI 10.1111/j.1466-8238.2006.00284.x)
- K. J. Gaston, « Global patterns in biodiversity », Nature, vol. 405, no 6783, , p. 220–227 (PMID 10821282, DOI 10.1038/35012228)
- Gaston, K. J., and T. M. Blackburn. 2000. Pattern and processes in macroecology. Blackwell Scientific, Oxford.
- B. A. Hawkins et J. A. F. Diniz-Filho, « The mid-domain effect cannot explain the diversity gradient of Nearctic birds », Global Ecology and Biogeography, vol. 11, no 5, , p. 419–426 (DOI 10.1046/j.1466-822x.2002.00299.x)
- L. N. Gillman, D. J. Keeling, H. A. Ross et S. D. Wright, « Latitude, elevation and the tempo of molecular evolution in mammals », Proceedings of the Royal Society B: Biological Sciences, vol. 276, no 1671, , p. 3353–3359 (PMID 19556254, PMCID 2817169, DOI 10.1098/rspb.2009.0674)
- B. A. Hawkins, J. A. F. Diniz-Filho et A. E. Weis, « The mid-domain effect and diversity gradients: is there anything to learn », American Naturalist, vol. 166, no 5, , E140–E143 (PMID 16224716, DOI 10.1086/491686)
- B. A. Hawkins et E. E. Porter, « Area and the latitudinal diversity gradient for terrestrial birds », Ecology Letters, vol. 4, no 6, , p. 595–601 (DOI 10.1046/j.1461-0248.2001.00271.x)
- H. Hillebrand, « On the generality of the latitudinal diversity gradient », The American Naturalist, vol. 163, no 2, , p. 192–211 (PMID 14970922, DOI 10.1086/381004, lire en ligne)
- W. Jetz et C. Rahbek, « Geometric constraints explain much of the species richness pattern in African birds », Proceedings of the National Academy of Sciences of the USA, vol. 98, no 10, , p. 5661–5666 (PMID 11344307, PMCID 33269, DOI 10.1073/pnas.091100998, Bibcode 2001PNAS...98.5661J)
- J. T. Kerr, M. Perring et D. J. Currie, « The missing Madagascan mid-domain effect », Ecology Letters, vol. 9, no 2, , p. 149–159 (PMID 16958880, DOI 10.1111/j.1461-0248.2005.00860.x)
- P. Koleff et K. J. Gaston, « Latitudinal gradients in diversity: real patterns and random models », Ecography, vol. 24, no 3, , p. 341–351 (DOI 10.1111/j.1600-0587.2001.tb00207.x)
- J. H. R. Lambers, J. S. Clark et B. Beckage, « Density-dependent mortality and the latitudinal gradient in species diversity », Nature, vol. 417, no 6890, , p. 732–735 (PMID 12066182, DOI 10.1038/nature00809, Bibcode 2002Natur.417..732H)
- D. C. Lees et R. K. Colwell, « A strong Madagascan rainforest MDE and no equatorward increase in species richness: Re-analysis of 'The missing Madagascan mid-domain effect', by Kerr J.T., Perring M. & Currie D.J (Ecology Letters 9:149-159, 2006) », Ecology Letters, vol. 10, no 9, , E4–E8 (PMID 17663706, DOI 10.1111/j.1461-0248.2007.01040.x)
- E. R. Pianka, « Latitudinal gradients in species diversity: a review of concepts », American Naturalist, vol. 100, no 910, , p. 33–46 (DOI 10.1086/282398)
- S.L. Pimm et J.H. Brown, « Domains of diversity », Science, vol. 304, no 5672, , p. 831–833 (PMID 15131295, DOI 10.1126/science.1095332)
- C. Rahbek, N. Gotelli, R.K. Colwell, G.L. Entsminger, T.F.L.V.B. Rangel et G.R. Graves, « Predicting continental-scale patterns of bird species richness with spatially explicit models », Proceedings of the Royal Society of London B, vol. 274, no 1607, , p. 165–174 (PMID 17148246, PMCID 1685854, DOI 10.1098/rspb.2006.3700)
- K. Rohde, « Latitudinal gradients in species diversity: the search for the primary cause », Oikos, vol. 65, no 3, , p. 514–527 (DOI 10.2307/3545569, JSTOR 3545569)
- K. Rohde, « The larger area of the tropics does not explain latitudinal gradients in species diversity », Oikos, vol. 79, no 1, , p. 169–172 (DOI 10.2307/3546102, JSTOR 3546102)
- J. Rolland, F.L. Condamine, F. Jiguet et H. Morlon, « Faster speciation and reduced extinction in the tropics contribute to the mammalian latitudinal diversity gradient », PLOS Biology, vol. 12, no 1, , e1001775 (PMID 24492316, PMCID 3904837, DOI 10.1371/journal.pbio.1001775)
- T. S. Romdal, R. K. Colwell et C. Rahbek, « The influence of band sum area, domain extent and range sizes on the latitudinal mid-domain effect », Ecology, vol. 86, , p. 235–244 (DOI 10.1890/04-0096)
- M. L. Rosenzweig, « Species diversity gradients: we know more and less than we thought », Journal of Mammalogy, vol. 73, no 4, , p. 715–730 (DOI 10.2307/1382191, JSTOR 1382191)
- Rosenzweig, M. L. 2003. How to reject the area hypothesis of latitudinal gradients. Pages 87–106 in T. M. Blackburn and K. J. Gaston, editors. Macroecology: Concepts and Consequences. Blackwell Publishing, Oxford.
- D. Storch, R.G. Davies, S. Zajicek, C.D.L. Orme, V. Olson, G.H. Thomas, T.S. Ding, P.C. Rasmussen, R.S. Ridgely, Tim M. Blackburn, Ian P. F. Owens et Kevin J. Gaston, « Energy, range dynamics and global species richness patterns: reconciling mid-domain effects and environmental determinants of avian diversity », Ecology Letters, vol. 9, no 12, , p. 1308–1320 (PMID 17118005, DOI 10.1111/j.1461-0248.2006.00984.x)
- J. Terborgh, « On the notion of favorableness in plant ecology », American Naturalist, vol. 107, no 956, , p. 481–501 (DOI 10.1086/282852)
- J.T. Weir et D. Schluter, « The latitudinal gradient in recent speciation and extinction rates of birds and mammals », Science, vol. 315, no 5818, , p. 1574–1576 (PMID 17363673, DOI 10.1126/science.1135590, Bibcode 2007Sci...315.1574W)
- M. R. Willig et S. K. Lyons, « An analytical model of latitudinal gradients of species richness with an empirical test for marsupials and bats in the New World », Oikos, vol. 81, no 1, , p. 93–98 (DOI 10.2307/3546471, JSTOR 3546471)
- M. R. Willig, D. M. Kaufmann et R. D. Stevens, « Latitudinal gradients of biodiversity: pattern, process, scale and synthesis », Annu. Rev. Ecol. Syst., vol. 34, , p. 273–309 (DOI 10.1146/annurev.ecolsys.34.012103.144032)
- S. Wright, J. Keeling et L. Gillman, « The road from Santa Rosalia: a faster tempo of evolution in tropical climates », Proceedings of the National Academy of Sciences, vol. 103, no 20, , p. 7718–7722 (PMID 16672371, PMCID 1472511, DOI 10.1073/pnas.0510383103, Bibcode 2006PNAS..103.7718W)
- F. A. Zapata, K. J. Gaston et S. L. Chown, « Mid-domain models of species richness gradients: assumptions, methods and evidence », Journal of Animal Ecology, vol. 72, no 4, , p. 677–690 (PMID 30893962, DOI 10.1046/j.1365-2656.2003.00741.x)
- F. A. Zapata, K. J. Gaston et S. L. Chown, « The mid-domain effect revisited », American Naturalist, vol. 166, no 5, , E144–E148 (PMID 16224717, DOI 10.1086/491685)
Voir aussi
- Biodiversité
- Biomes
- Temps évolutif efficace (en)
- Évolution
- Extinction
- Latitude
- Spéciation
- Richesse spécifique
- Tropique