Contrainte (biologie)
La contrainte est un concept central en Évolution et développement, dans la discipline de la biologie. Elle permet de relier l’Évolution et le développement. Elle s’oppose à la sélection pour certains et pour d’autres elles sont en interaction. La contrainte essaie de répondre à la question qui de l’Évolution ou du Développement précède l’autre en alliant les deux de manière qu’elle ne puisse être l’une sans l’autre.
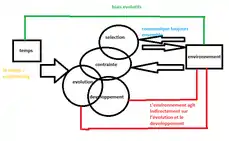
La contrainte en écologie évolutive est une entrave à une évolution dans certaines directions ou empêchant totalement certaines trajectoires évolutives[1].

Ces contraintes peuvent être réparties en 4 catégories : génétiques, sélectives, fonctionnelles et développementales. Les contraintes fonctionnelles (et développementales) sont les plus importantes puisque ce sont elles qui conditionnent les autres contraintes. Ces contraintes sont liées aux lois de la physique, au temps ou encore à l’énergie et sont donc à l’origine de compromis évolutifs ou trade-offs[2].
Évolution-Développement en biologie
Cette discipline émergente est à la croisée de toutes les notions en biologie, que l'on nomme Evo-Devo dans le jargon biologique[3]. Elle se revendique très jeune, avec une approche très large de ce que pourrait être l'évolution tant dans la biomoléculaire que dans la génétique, l’écologie, etc. Le développement apporte des biais ou des changements génétiques qui peuvent changer le rythme et la direction de l’Évolution, et cela par l’intermédiaire de la contrainte. L’Évolution est quant à elle très intrinsèquement liée au temps. La notion de temps est un concept clé dans cette discipline car elle un sens à l’existence de tous les questionnements posés par Evo-Devo[4].
Contrainte et sélection
Une contrainte peut être vue comme un choix, ou comme le fait d'imposer un choix[5], entre ces deux approches, deux théories se font face.
La contrainte donne une direction, et de fait impose une voie par rapport à une autre, ou peut englober plusieurs choix par rapport à d'autres, une contrainte enfin, peut être vue comme existante car relative à une autre. La sélection découle d’un processus de compétition pour une adaptation toujours plus optimale. Deux théories se font face : la première opposant la contrainte à la sélection et la deuxième assume que l’une ne va pas sans l’autre, et qu’elles sont inter reliées, prônée en Evo-Devo.
Selon la deuxième théorie, la contrainte se manifeste par la sélection. Et la sélection existe de par la contrainte. En d’autres mots, les contraintes peuvent être vues comme les délimitations de voies de changements et la pression sélective comme la force générant ce changement[6].
La contrainte comme cadre d'analyse
En parlant de ce sujet, resituer à chaque niveau d’analyse la contrainte par rapport à une autre, permet de comprendre les mécanismes de patterns sous-jacents, exposé plus bas, pour saisir la pleine dimension que la notion de contrainte peut évoquer dans une discipline. On peut parler de contrainte relatives, absolue, générative, mais aussi de contraintes développementales ou évolutives, de contraintes en Éco-Devo. Ce qui limite les contraintes de manières fonctionnelles sont le temps, l’énergie et les lois physiques. Une contrainte en influence une autre. Une contrainte est générée par une contrainte. Et cela peut se répéter dans un pattern infini, tout dépend de jusqu’où l’on part… du big bang à la molécule! On comprend que la dissociation d’une contrainte par rapport à une autre varie en fonction du sujet et de l’échelle d’analyse. Ainsi une contrainte pourrait Être, exister, et donc se justifier par le fait de la pression qu’une autre contrainte venant d’un autre cadre d’analyse ou d’un autre environnement viendrait appuyer appuyer sur celle-ci[7].
Un exemple : une contrainte phylogénétique, avec un point d’origine différent entre deux espèces est encadrée par une contrainte Eco-Devo, où, en raison de l’évolution convergente des niches (dues certainement aux conditions environnementales et écologiques) nous font aujourd’hui confondre ces animaux dans leur terminologie. On doit donc faire attention à comprendre les processus de l’évolution, et la contrainte peut être un bon fil d’Ariane, avec ses limites de données à elle aussi.
Le labyrinthe : une métaphore de la contrainte
Le labyrinthe est un exemple très parlant de tous ces mécanismes. Il peut aider à mettre une image sur cette notion qu’est la contrainte. L’Évolution serait l’entité perdue dans le labyrinthe, représenté par le point rouge dans les figures 1(la sélection va vers quelque chose, est animée d'un développement), 2 (Une contrainte pose un choix, qui se décide de par la sélection), 3 (Labyrinthe montrant un chemin Évolutif orienté par la sélection). Chaque mur du labyrinthe est une contrainte et le développement est représenté par la flèche rouge, dans la figure 1, c’est le point de départ à la suite d'une première contrainte. Dans la figure 2, on se retrouve devant un choix face à une contrainte. La sélection détermine quel est le mécanisme adaptatif optimal et ceci dans un but de trouver la sortie la plus proche comme Climax, équilibre entre environnement et l’individu, par exemple, en Evo-Devo. On le voit dans la figure 3, où l’Évolution serait le tracé vert. Les « détours » possibles n’ont pas été représentés pour ne pas surcharger le schéma. Le point d’interrogation suggère qu’il n’y a pas de bonne ou mauvaise évolution, et souligne le caractère résolument adaptatif de l’évolution, à travers différents processus de développement générés par les contraintes et les processus de sélection. L’autre flèche rouge sous-entend qu’il peut y avoir des biais dans les voies adoptés, on peut prendre plusieurs chemin peuvent amener à une sortie, et si l’on considère une sortie en particulier par rapport à une autre, alors qu’elles se ressemblent toute (il est difficile de savoir combien il y a de sortie dans un labyrinthe et quelle sortie est différente de l’autre) on pourrait se leurrer, et donc cela illustre les biais en développement, ou encore les changements ou innovations génétique. La contrainte est appliquée à la sélection[8]. Ici, article propose une approche entre l'espace et la contrainte d'un point de vue de l'urbanisme, où comment l'homme progresse dans son environnement.
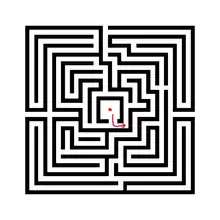


La question clé sur la direction de l'Évolution
La sélection naturelle, au niveau des populations et le développement au niveau inter et intra spécifique, sont parmi les facteurs qui guident le sens de l’Évolution.
Paysage adaptatif
Cet exemple est inspiré du modèle de paysage adaptatif introduit par Sewall Wright. On modélise la contrainte comme chemin emprunté par l’Évolution et orienté par le développement. Le chemin tend vers des vallées qui sont le point de climax et d’équilibre. Celui-ci est d’ailleurs toujours en perpétuel mouvement, car inhérent à la variable temps. Ce point de climax est une image factice vers laquelle l’évolution tend en s’adaptant toujours de manière à son environnement et aux contraintes imposées par cet environnement. Le chemin est donc la contrainte ou encore, en d’autres mot, e vecteur ou la voie de changement du développement par un processus de sélection. À travers ce modèle on peut présenter brièvement les 4 grands types de contraintes qui reviennent en Evo-Devo :
- La contrainte absolue : où le fait même de l’existence du chemin est par le fait d’un ensemble de contraintes qui agissent sur ce chemin[9].
- La contrainte relative : une contrainte existe par rapport à une autre contrainte d’un même point focal. Les deux chemins sont des contraintes relative l’une à l’autre[9].
- La contrainte en Eco-Devo : Rassemble l’ensemble des facteurs environnementaux et écologiques qui pourraient s’illustrer ici par la texture du chemin versus la bordure autour de ce chemin. On note un changement sur les lois physiques[9].
- La contrainte développementale : subit les biais et changements génétique. Cette contrainte peut s’opposer à une contrainte d’un autre type comme une contrainte de l’environnement. On assiste à un effet de pression de changements évolutifs de l’environnement sur un individu par exemple.
- La contrainte Évolutive : c’est la contrainte associée au processus de l’évolution. Elle encadre cette Évolution, la bordure du chemin dans l’exemple du paysage[9].
La contrainte peut se voir comme un support, une structure donnant forme aux choix évolutifs, aux changements génétiques. Elle peut également se comprendre comme assujettie à une notion de mouvement, avec l'Évolution qui apporte a dimension temporelle.
La contrainte comme variable limitante
La contrainte peut être aussi un mécanisme ou processus limitant la variabilité infinie de l’évolution de caractères, au moment de la création de Vie. Mais aussi au fil des générations au sein d’une population. La contrainte peut biaiser ou limiter la réponse évolutive. Par des interférences venant des processus de sélection par exemple. En effet, sélection et contrainte agissent l’une envers l’autre. Ainsi, l’observation au niveau des phénotypes peut être biaisée, car on n’est pas capable de remonter e processus de contrainte en question. On ne peut connaitre exactement les conditions environnementales et la période temporelle exacte où l’événement a eu lieu, pour définir la genèse de cette contrainte et sa causalité. En outre, c’est dans l’environnement que nous comprenons les processus de sélection au sein d’une population. Le mécanisme de sélection permet aux plus adaptés de survivre et de maintenir le fitness de l’espèce. Une des causalités de la contrainte est d’affecter la réponse évolutive d’un trait due à des facteurs de sélection extérieurs. La majorité des contraintes agissent à un stade précoce de développement pour contrôler à variabilité génotypique, afin de maintenir un phénotype particulier, qui est sélectionné pour se maintenir au sein de la population. La contrainte, en fonction du point de focal, peut avoir comme but de limiter les changements génétiques que la pression de sélection apporte à un trait génétique par exemple. La contrainte maintient le génotype de l’espèce ou de l’individu face à d’autres contraintes extérieures.
Le temps, les patterns et la contrainte
Les contraintes interagissent avec d’autres contraintes. Mais on observe également un pattern de contraintes qui se maintiennent plus ou moins dans le temps. Comme une contrainte peut en encadrer une autre, on peut supposer qu’il en est de même pour les patterns de contraintes retrouvés dans la Nature. Un pattern consiste en une répétition de même schéma à l’infini. Cela sous-tend à une certaine dynamique, un rythme dans les apparitions de ces contraintes. Ces fréquences de contraintes sont très certainement liées aux variations génétiques, phénotypiques et aux innovations génotypiques également.
Un des gros débats en Evo-Devo, est à propos de la raison des variations dans le génome observés au cours de l’Évolution. On pose véritablement la question de qui est venu en premier de l’œuf ou la poule. Comment apparaissent les innovations génétiques, est-ce que ce sont les variations génétiques qui provoquent les changements phénotypiques à cause d’une pression de sélection ou est-ce déjà programmé ? Ou mathématiquement prévisible ? Peu d’analyses sont encore faites à ce sujet.
On peut supposer qu’il y a une sorte de ratio entre des contraintes fixes et des contraintes aléatoires, et qu’ils s’alternent à une certaine fréquence et redondance dans le temps. L’alternance de nouvelles contraintes dans un pattern de contraintes déjà fixées pourrait être à l’origine d’innovations ou changements génétiques. Ainsi on perd l’équilibre, le point de climax se déplace à nouveau vers un autre équilibre et provoque de fait une cascade de variations dans l’environnement impliquant une nouvelle dynamique de sélection et l’apparition de nouvelles contraintes. Par exemple, le changement de taille des pinces de crabes, elles furent un moment plus grandes du côté droit par rapport au côté gauche, puis à un moment précis, ce fut l’inverse. On appelle cela une allométrie phénotypique. Le mystère est ici de connaitre les raisons de ce soudain changement phénotypique, cela passe par la connaissance de l’environnement à cette période de changement. La logique évolutive semble tendre vers une logique où certains changements ont une valeur adaptative nettement plus efficace que d’autres et se conservent dans le temps. Les patterns se répètent par rapport à ces traits que l’on veut conserver et tout le mécanisme de l’évolution repart, dans ce cercle, parsemé de fractures, innovations, et nouveaux cycles. La contrainte suit ce même rythme, à une échelle autant macroscopique que microscopique, individuelle ou populationnelle.
Nous retrouvons la contrainte dans beaucoup de cadres et notions d'Évo-Devo, elle peut s’impliquer dans chaque exemple de la discipline. Cette notion est large et plutôt abstraite, mais elle est partie de la pierre angulaire de cette discipline qu’est l'Evo-Devo[10].
Trade-offs en écologie évolutive
En écologie évolutive, un trade-off (ou compromis évolutif) correspond à une situation dans laquelle le changement d'un trait associé à une meilleure fitness implique en retour la modification d'un autre trait qui est lui associé à une baisse de fitness[1].
Ces trade-offs peuvent être observés tant à l'échelle intra qu'interspécifique.
Il existe 3 niveaux de trade-off : phénotypique, intermédiaire et génotypique. Au niveau phénotypique, on s’intéresse à des traits liés directement à la survie et à la reproduction. C’est ce niveau qui explique que les organismes n’aient pas une fécondité extrêmement haute. Le niveau génétique se rapporte aux traits induits par la génétique. Le niveau intermédiaire constitue tout ce qui fait le lien entre génotype et phénotype, avec notamment le contrôle hormonal d’allocation de ressources pour chaque fonction[1].
Énergie allouée et modèle en Y
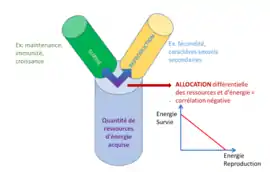
Il est facile d'imaginer qu’un individu maximiserait sa fitness en ayant à la fois un très fort succès reproducteur et une grande capacité de survie. L’absence de ce type d’individu dans la nature est expliquée par le modèle en Y[11], qui se base sur le principe que la quantité de ressources (et donc d'énergie) acquise par l'individu est finie et limitée. Ainsi, la quantité allouée à une certaine fonction sera alors indisponible pour les autres fonctions. Afin d'illustrer ce propos, il est possible de séparer les fonctions en deux types : celles qui contribuent à la reproduction (fécondité, investissement dans les structures sexuelles…) et celles qui contribuent à la survie (maintenance, croissance…). Ces deux catégories sont les composantes de la fitness. L'énergie allouée dans une fonction n’étant pas investie pour l’autre, on parle alors de fonctions antagonistes.
On peut définir la relation suivante T = X1 + X2. Où X1 et X2 sont des traits déterminés par l'allocation des ressources d'un pool commun, T[12].
Un compromis évolutif se traduit dans les cas simples par une corrélation négative entre deux ou plusieurs traits (voir image).
Évolution des trade-offs
S'il existe un trade-off entre deux fonctions, il est facile de penser qu’il puisse exister un compromis optimal entre ces deux fonctions (c'est-à-dire un compromis qui maximise la fitness des individus). Or, dans la plupart des cas, on observe encore une variabilité dans la Nature. Cette hétérogénéité peut être expliquée par quatre hypothèses :
- Dans un premier temps, la variabilité peut être expliquée par l’équilibre mutation-sélection[13]. En effet, de nouvelles mutations peuvent modifier le compromis optimal et donc expliquer les divergences observables.
- La variation peut être également expliquée par la « pléiotropie antagoniste »[14], c'est-à-dire lorsque certains gènes sont impliqués dans plusieurs fonctions. Par conséquent, un gène augmentant la fitness d’un individu en améliorant une fonction peut également avoir un impact négatif sur une deuxième fonction et de ce fait diminuer la fitness cet individu.
- De plus, l’absence de compromis optimal peut être dû à une sélection corrélationnelle : lorsque la sélection favorise la combinaison de certains traits. De ce fait, plusieurs combinaisons de traits différents sont observables s’ils donnent lieu à une fitness similaire.
- Enfin, cela peut tout simplement être une conséquence de la variabilité de l’environnement. En effet, une variabilité environnementale donne lieu à différentes pressions de sélection et donc à une variation des combinaisons optimales[1].
Influence des habitats sur les compromis évolutifs
La classification de Richard Levins (1968), permet de distinguer deux grands types d’habitats pouvant influencer les compromis évolutifs.
- Habitats à fort coût de reproduction :
Dans les habitats à fort coût de reproduction, la fitness des individus dépend majoritairement de leur taille. En effet, dans ces environnements, les plus grands individus sont les meilleurs compétiteurs, ils ont une meilleure survie et donc une meilleure fitness. Les caractéristiques de ces habitats vont alors mener à des individus investissant plus de ressources et d'énergie dans les traits permettant une forte croissance et de ce fait investissant moins de ressources dans les traits concernant la reproduction.
- Habitats à faible coût de reproduction :
Dans les habitats à faible coût de reproduction, la fitness des individus ne dépend que très peu de leur taille. Cela peut être le cas dans les environnements où il y a une absence de prédation, de concurrence ou encore une mortalité préférentielle pour les grands individus. Ces types d’habitats favorisent donc les individus investissant plus d'énergie dans la reproduction que dans la croissance[15].
Observation des compromis évolutifs
L’observation de corrélations négatives démontrant la présence de compromis évolutifs peut être difficile dans les conditions réelles[15]. En effet, le manque de variation entre individus au sein d’une population peut empêcher d’observer une corrélation négative . Au contraire, s’il y a de trop fortes différences, les individus possédant la meilleure fitness vont pouvoir investir dans tous les traits. Dans cette situation, une corrélation positive peut alors être observée. De plus, il est possible que les deux traits étudiés soient dépendants d’un troisième[16]. En effet, il est possible d’observer une corrélation positive si par exemple une mauvaise condition physique réduit le taux de survie ainsi que la fécondité. Autrement dit, il existe de nombreux biais susceptibles d’induire en erreur l’étude des compromis évolutifs.
Afin de mettre en évidence un compromis évolutif, plusieurs types d’approches sont possibles.
- Approche génétique
Il est possible d’étudier à travers une étude expérimentale des liaisons génétiques (et donc phénotypiques) entre plusieurs traits[1]. Ces expériences se font en sélectionnant des lignées génétiques ayant différents niveaux pour un trait pour ensuite regarder la variation d'un deuxième trait pour ces mêmes lignées. Le trade-off est mis en évidence par l'obtention d'une corrélation négative entre ces deux traits (dont on sait qu'ils dépendent d'un contrôle génétique[17]). Il est néanmoins assez difficile d'exploiter des résultats par approche génétique, notamment par le fait que les approches corrélatives permettent juste de montrer un lien entre ces deux traits mais pas d'en expliquer les causes.
- Contrôle expérimental des ressources
Il est possible que la corrélation entre deux traits dépende de l’environnement. En effet, chez les populations femelles de charançon Callosobruchus maculatus, on observe une corrélation positive entre la fécondité et la survie dans des conditions réelles (en effet, mieux un individu survit, plus il aura de descendants). Cependant, dans des conditions expérimentales où l’on limite l’accès à la nourriture, on observe que la fécondité moyenne est réduite mais que la survie augmente. Un trade-off peut donc être invisible dans les conditions réelles mais apparaître dans des conditions expérimentales avec une manipulation de l’accès aux ressources[18].
Implications dans la gestion des ressources marines
Les femelles morues de l’Atlantique (Gadus morhua) atteignent leur maturité sexuelle à partir de 60cm. Elles pondent environ 2 millions d'œufs par an à 80 cm mais pondent jusqu’à 15 millions d'œufs à 120cm. Ces chiffres illustrent le trade-off entre croissance et reproduction. En effet, chez la plupart des organismes à croissance continue, il existe un compromis entre âge et taille à la reproduction (compromis entre se reproduire tôt avec peu de descendants ou plus tard avec plus de descendants). L’industrie de la pêche a exploité les plus grands (et donc les plus vieux individus) or ceux-ci représentaient les meilleurs reproducteurs de la population. Cette pression de sélection a induit une baisse significative de la taille et de l’âge à maturité des morues conduisant donc, à une baisse de la quantité totale d'œufs pondus due au trade-off[19].
Quelques exemples de trade-offs
Beaucoup de trade-offs concernent le coût de la reproduction et s’expriment par une diminution de la survie des parents, ou par un compromis entre la taille et le nombre de descendants. Il est possible de définir trois grandes catégories de trade-off:
- Exemple de trade-off reproduction/survie
Les anoles bruns(Anolis sagrei) sont des reptiles qui ne pondent qu'un œuf par an. Il a été démontré que les individus ayant subi une chirurgie de stérilisation possédaient une meilleure survie que les individus pouvant se reproduire. Ceci met en évidence un trade-off reproduction/survie, et montre qu'une énergie importante est utilisée pour la reproduction (même lorsque seul un œuf par an est pondu). Cette énergie qui n'est alors pas utilisée pour d'autres fonctions impliquant la survie de l'individu[20].
- Exemple trade-off nombre de descendants/taille des descendants
Il a été montré chez plusieurs espèces d'anatidés, qu'il existe une corrélation entre la taille et le nombre d’œufs. En effet, plus les œufs sont gros, moins ils sont nombreux. Ceci met en évidence un trade-off interspécifique, entre taille de la descendance et nombre de descendants[21]. On peut mettre ceci en lien avec les stratégies de reproduction R et K, selon laquelle les espèces ont soit beaucoup de petits descendants (stratégie R) soit peu de grands descendants (stratégie K).
- Exemple trade-off énergie allouée à la reproduction/fitness des descendants
On parle de trade-off intergénérationnel lorsque l’énergie allouée par la femelle pour la reproduction joue un rôle sur la fitness de ses descendants[1]. Il a par exemple été montré que le nombre d'œufs pondus ne constituait pas uniquement un compromis avec la taille des œufs mais également avec les chances de reproduction des descendants[22].
Voir aussi
Bibliographie
- (en) Paul M. Brakefied, Evo-devo and constraints on selection, Leiden University, Netherlands, TRENDS in Ecology and Evolution, , 362-367 p.
- (en) Paul M. Brakefied, The power of evo-devo to explore evolutionary constraints : experiments with butterfly eyespots, Evolution and development, , 283-290 p.
- (en) Giuseppe Fusco, How many processes are responsible for phenotypic evolution, Evolution and development, , 279-286 p.
- (en) Paul M. Brakefield et J.C Roskam, Exploring Evolutionary Constraints Is a Task for an Integrative Evolutionary Biology, The American Naturalist, , S4-S13
- (en) Stevan J. arnold, Constraints on Phenotypic Evolution, The American Naturalist, , S85-s107
- (en) Wendy M. Olson et Brian K. Hall, Keywords and concepts in evolutionary developmental biology, Cambridge (Mass.), Harvard University Press, , 476 p. (ISBN 0-674-00904-5)
- (en) Wallace Arthur, A Theory of the Evolution of Development, Chichester/New York/Brisbane etc., John Wiley & Sons Canada, , 94 p. (ISBN 0-471-91974-8)
Articles connexes
Notes et références
- S. C. Stearns, « Trade-Offs in Life-History Evolution », Functional Ecology, vol. 3, no 3, , p. 259–268 (ISSN 0269-8463, DOI 10.2307/2389364, lire en ligne, consulté le )
- Stevan J. Arnold, « Constraints on Phenotypic Evolution », The American Naturalist, vol. 140, , S85–S107 (ISSN 0003-0147, DOI 10.1086/285398, lire en ligne, consulté le )
- (en) Wallace Arthur, A Theory of the Evolution of Development, , 94 p. (ISBN 978-0-471-91974-2, lire en ligne).
- (en) J C Roskam, « Exploring evolutionary constraints is a task for an integrative evolutionary biology - PubMed », The American naturalist, vol. 168 Suppl 6, , S4–S13 (ISSN 1537-5323, PMID 17109328, DOI 10.1086/509049, lire en ligne, consulté le ).
- « implications-philosophiques.or… »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?).
- (en) Paul M Brakefield, « The power of evo-devo to explore evolutionary constraints: experiments with butterfly eyespots - PubMed », Zoology (Jena, Germany), vol. 106, no 4, , p. 283–290 (ISSN 0944-2006, PMID 16351914, DOI 10.1078/0944-2006-00124, lire en ligne, consulté le ).
- (en) « Developmental Constraints and Evolution : A Perspective from the Mountain Lake… », sur JSTOR (consulté le ).
- Liégeois, Laurence, « Espace labyrinthique et contrainte », sur revues.org, Géographie et cultures, Laboratoire Espaces, Nature et Culture (ENEC), (ISBN 978-2-296-10874-5, ISSN 1165-0354, consulté le ), p. 37–56.
- PM Brakefield, « Evo-devo and constraints on selection », Trends Ecol Evol., vol. 21, no 7), , p. 362-368 (PMID 16713653).
- (en) « Keywords and Concepts in Evolutionary Developmental Biology — Brian K. Hall, Wendy M. Olson », sur harvard.edu (consulté le ).
- G. de Jong et A. J. van Noordwijk, « Acquisition and Allocation of Resources: Genetic (CO) Variances, Selection, and Life Histories », The American Naturalist, vol. 139, no 4, , p. 749–770 (ISSN 0003-0147 et 1537-5323, DOI 10.1086/285356, lire en ligne, consulté le )
- Frédéric,. Thomas, Thierry,. Lefevre et Michel,. Raymond, Biologie évolutive, De Boeck supérieur, dl 2016, cop. 2016 (ISBN 978-2-8073-0296-9 et 2-8073-0296-3, OCLC 946571046, lire en ligne), p. 352, 370
- (en) Michael Lynch, « Evolution of the mutation rate », Trends in Genetics, vol. 26, no 8, , p. 345–352 (DOI 10.1016/j.tig.2010.05.003, lire en ligne, consulté le )
- James W. Curtsinger, Philip M. Service et Timothy Prout, « Antagonistic Pleiotropy, Reversal of Dominance, and Genetic Polymorphism », The American Naturalist, vol. 144, no 2, , p. 210–228 (ISSN 0003-0147 et 1537-5323, DOI 10.1086/285671, lire en ligne, consulté le )
- Claire Tirard, Luc Abbadie, David Laloi et Philippe Koubbi, Ecologie - Fiches de cours, exemples appliqués et QCM, (lire en ligne), Partie 1, Chapitre 3: Histoire de vie, compromis, stratégies d’allocation.p82, p79
- Claude Henry, Biologie des populations animales et végétales, (lire en ligne), Partie 2, chapitre 8: Génétique, sélection et évolution des traits de vie quantitatifs. p492
- Derek A Roff et Michael J Bradford, « Quantitative genetics of the trade-off between fecundity and wing dimorphism in the cricket Allonemobius socius », Heredity, vol. 76, no 2, , p. 178–185 (ISSN 0018-067X et 1365-2540, DOI 10.1038/hdy.1996.25, lire en ligne, consulté le )
- (en) Frank J. Messina et Andrea F. Slade, « Expression of a life-history trade-off in a seed beetle depends on environmental context », Physiological Entomology, vol. 24, no 4, , p. 358–363 (ISSN 1365-3032, DOI 10.1046/j.1365-3032.1999.00151.x, lire en ligne, consulté le )
- Michael L. Cain, Ecology, Sinauer Associates, (ISBN 978-0-87893-083-8 et 0-87893-083-3, OCLC 196314782, lire en ligne), p. 166
- Robert M. Cox et Ryan Calsbeek, « Severe costs of reproduction persist in Anolis lizards despite the evolution of a single-egg clutch », Evolution; International Journal of Organic Evolution, vol. 64, no 5, , p. 1321–1330 (ISSN 1558-5646, PMID 19930451, DOI 10.1111/j.1558-5646.2009.00906.x, lire en ligne, consulté le )
- (en) David Lack, « The significance of clutch-size in waterfowl », Wildfowl, vol. 18, no 18, , p. 125–128 (ISSN 2052-6458, lire en ligne, consulté le )
- M. Mangel, J. A. Rosenheim et F. R. Adler, « Clutch size, offspring performance, and intergenerational fitness », Behavioral Ecology, vol. 5, no 4, , p. 412–417 (ISSN 1045-2249 et 1465-7279, DOI 10.1093/beheco/5.4.412, lire en ligne, consulté le )