Sélection fluctuante
La sélection fluctuante est une des composantes de la sélection naturelle, où l'on observe des variations d'intensité et de direction de la sélection[1]. Ces variations entraînent la fluctuation d'allèles et donc l'alternance de phénotypes au sein d'une population dans un temps relativement court à l'échelle de l'évolution. Darwin[2] a défini la sélection comme étant principalement directionnelle et un processus lent de changements évolutifs[3]. Cependant Stephen Jay Gould a mis en évidence qu'il existe des exemples d'évolution rapide, comme dans le registre fossile où l'on retrouve des étapes d'alternance de diversification instantanée et d'état de stase, c'est la sélection fluctuante.
Définition
Dans le cadre de la sélection fluctuante, l'évolution des caractères n'est pas linéaire. La sélection d'allèle dans une population peut passer par des phases d'intensités et de directions différentes:
- L'intensité de la sélection est une composante du taux d'évolution : Taux d'évolution = vitesse d'apparition x intensité de sélection. Cette intensité est guidée par les variations (plus ou moins forte) de l'environnement, qui font pression sur la sélection.
- la direction de la sélection correspond à l'orientation que va prendre l'évolution pour un caractère. Des allèles différents peuvent apparaître de manière simultanée pour un même locus dans une population.
Ces deux propriétés sont en majorité régulés par l'environnement. Il en découle un état fondamental de la sélection fluctuante, la diversité allélique. La fluctuation d'intensité et de direction de la sélection entraîne l'apparition d'allèles sur même locus dans une population. L'abondance de ces allèles va fluctuer au sein de la population au cours du temps et ces allèles vont être maintenues sans entraîner la sélection définitive de l'un des allèles.
Variations environnementales
Les variations de la sélection fluctuante sont aussi fortement liées aux variations environnementales[4]. Le changement de direction et d'intensité de la sélection est fortement guidé par les fluctuations environnementales qui peuvent survenir à plus ou moins court termes. Dans une population donnée, les conditions environnementales du milieu (température, précipitation, profondeur de l'eau) entraînent la sélection de caractère adapté à ce milieu. Ces caractères se fixent dans la population et deviennent héritables entre les générations. Lorsque les conditions environnementales fluctuent, les individus de la population sont obligés de s'adapter en sélectionnant de nouveaux caractères[5]. Cependant, si les fluctuations environnementales sont de courte durée, le nouveau caractère n'a pas le temps de se fixer dans la population et les deux variations alléliques sont conservées. Lorsque les conditions environnementales changent rapidement, la proportion de l'allèle la plus adaptée sans entraîner la disparition de l'autre allèle. La sélection fluctuante permet, par les variations de directions de la sélection, de conserver au sein d'une population plusieurs stratégies.
Reine Rouge et interactions hôtes-parasites
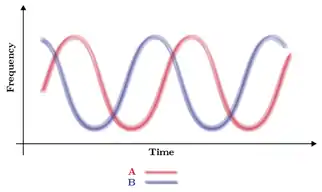
L'hypothèse de la reine rouge est à l'origine de stratégies de compétition évolutive interspécifique. Dans le cas des interactions hôtes-parasites, par exemple, elle se base sur une compétition interspécifique où chaque espèce a pour but de contourner les stratégies offensives et défensives de son antagoniste.
Dans ce cas, la vitesse d'évolution dans la course à l'armement est très rapide au début de leur interaction car leur relation n'est pas spécifique et qu'il existe des failles exploitables dans leurs stratégies. Mais à partir d'un certain temps, la coévolution entre les espèces, se spécialisant de plus en plus les unes par rapport aux autres, s'essouffle. Il y a maintien dans la population d'hôtes et de parasites de plusieurs phénotypes différents qui seront fréquence dépendant. En effet, une stratégie généraliste est coûteuse car elle demande d'être adaptée dans plusieurs domaines. La sélection fluctuante permet de garder dans la population d'hôte plusieurs allèles différents qui vont s'exprimer en plus ou moins grande quantité en fonction de la présence ou non des parasites. Par exemple, dans le cas d'une possible alternative entre l'apparition d'un parasite A et d'un parasite B : si un parasite A apparaît, alors la proportion d'hôte sera majoritairement des hôtes A avec des résistances adaptées à ce type de parasite. Les hôtes B, avec des résistances moins ou non adaptées aux parasites A seront minoritaires. Ces proportions s'inversant lors de l'apparition de parasite B entraînant ainsi le maintien à long terme de ces deux phénotypes. Suit on suit la fréquence des allèles A et B au cours du temps on remarque une fluctuation de l'abondance (Figure).
La sélection fluctuante permet de stabiliser la dynamique de course à l'armement lorsque ces derniers ne décident pas de collaborer[6]. La course au armement s’arrête car trop coûteuse en énergie, et la variabilité phénotypique se stabilise chez les deux espèces, cet événement est observable lorsqu'il n'y a pas de mutualisme.

Exemple de Diaptomus sanguineus
Les fluctuations dans les stratégies de ponte de copépodes dans les lacs d'eau douce est en majorité liée aux variations environnementales.
La date d'éclosion des œufs de copépodes d'eau douce a lieu au printemps, et peut être modifiée par des paramètres physico-chimiques. Lorsque la hauteur de la colonne d'eau est élevée, il y a une forte dilution du risque de prédation, et inversement. Les poissons arrivent au printemps pour frayer, ce qui augmente la pression de prédation sur les copépodes d'autant plus si le niveau de l'eau est bas. Au moment de la ponte, les copépodes peuvent induire une naissance immédiate des œufs ou bien un phénomène de diapause (retard de l’éclosion). La diapause recule l'éclosion des œufs de copépodes qui permet de survivre à la période du frai des poissons. Cependant cette stratégie entraîne une diminution de fitness car les copépodes pondent dès l’éclosion, ils pondent donc moins d’œufs que leurs congénères. Hairston[7] a réussi à mettre en évidence que ce comportement était héritable et devrait entraîner la mise en place d'une SES. Mais Hairston Junior[8] a montré qu'à chaque événement saisonnier d'éclosion, la fréquence des deux différentes stratégies était soit proche de 0 ou bien de 1. L’intégralité de la population choisit soit l'une ou l'autre des stratégies d'une saison sur l'autre. Or, cela n'entraîne pas la sélection d'une stratégie à long terme, il y a bien fluctuation de deux formes alléliques.
Le caractère fluctuant de l'environnement n'entraîne pas la sélection d'une stratégie, ou bien la sélection de deux stratégies (sélection disruptive) mais le maintien de deux caractères héritables par ce qui semble être de la sélection fluctuante. D'une saison sur l'autre, les deux stratégies survivent mais la proportion de l'une ou l'autre est guidée par les fluctuations environnementales.
Conséquence
La sélection fluctuante induit des bouleversements dans la compréhension des adaptations. La théorie synthétique de l'évolution définit un nouveau concept dans la compréhension de l'évolution des caractères, le paysage adaptatif. Il décrit la relation entre un caractère et sa fitness. La sélection devrait pousser l'évolution des caractères vers une valeur unique donnant une fitness optimale. Cependant cet optimum n'est valable que lorsque l'environnement est stable, car le paysage adaptatif est fixe. D'après la sélection fluctuante l'environnement n'est pas stable, le paysage adaptatif ne reste donc pas fixe, l’optimum du caractère va sans cesse se déplacer. La sélection fluctuante impose donc une nouvelle règle au concept d'adaptation, les variations environnementales rendent donc impossible une adaptation totale d'un individu à long terme[9].Il ne s'agit pas d'un cas isolé de sélection, mais bien du cas générale de la sélection naturelle, c'est au contraire la sélection directionnelle qui est un cas particulier.
Notes et références
- David, M. (2011). Personnalité, stratégies d'approvisionnement et d'appariement chez les Diamants Mandarins (taeniopygia guttata) (Doctoral dissertation, Université de Bourgogne; Université du Québec à Montréal).
- Darwin, C. (1965). La Selection naturelle.
- Bell, G. (2010). Fluctuating selection: the perpetual renewal of adaptation in variable environments. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 365(1537), 87-97.
- Lin, W. H., Rocco, M. J., Bertozzi‐Villa, A., & Kussell, E. (2015). Populations adapt to fluctuating selection using derived and ancestral allelic diversity. Evolution.
- Cain, A. J., Cook, L. M., & Currey, J. D. (1990). Population size and morph frequency in a long-term study of Cepaea nemoralis. Proceedings of the Royal Society of London B: Biological Sciences, 240(1298), 231-250.
- Hall, A. R., Scanlan, P. D., Morgan, A. D., & Buckling, A. (2011). Host–parasite coevolutionary arms races give way to fluctuating selection. Ecology letters, 14(7), 635-642.
- Hairston, N. G., & Olds, E. J. (1984). Population differences in the timing of diapause: adaptation in a spatially heterogeneous environment. Oecologia, 61(1), 42-48.
- Hairston Jr, N. G., & Dillon, T. A. (1990). Fluctuating selection and response in a population of freshwater copepods. Evolution, 1796-1805.
- Gould, S. J. (2006). La structure de la théorie de l'évolution. Gallimard.