Développement à régulation
Le développement à régulation est une théorie qui fait partie de la biologie évolutive du développement. Elle est souvent mise en opposition avec une autre théorie développementale, celle du développement en mosaïque.
Le principe de la théorie du développement à régulation
Ses expériences à l'appui, Hans Driesch a postulé en 1909 que les cellules embryonnaires peuvent s'adapter à tout changement qui éloigne l'embryon de son développement normal telles que la délétion, le déplacement ou l'ajout d'un blastomère. Il y aurait donc un mécanisme permettant de faire la régulation des blastomères pour s'assurer que l'embryon arrive à se développer correctement malgré les changements qu'il peut subir. Sans l'appui de la génétique, Hans Driesch a avancé que les cellules ont un potentiel de totipotence, c'est-à-dire qu'elles possèdent la capacité de répondre aux besoins du reste de l'embryon pour être en mesure de former un individu normal. Les cellules seraient donc capables de régénérer le contenu des cellules qui ont été enlevées[1].
L'histoire menant à la théorie du développement à régulation
L'embryologie est une science qui a suscité plusieurs controverses au fil du temps. Cette science a été étudiée et développée à partir des années 1850 et pendant une centaine d'années sans aucune considération pour la génétique. Les gènes n'étaient donc pas pris en compte dans le développement des embryons. Des scientifiques tels que Hans Driesch, Sven Hörstadius et Hans Spemann ont quand même travaillé très fort pour faire avancer cette science sans l'apport de la génétique et la compréhension de la force des gènes dans le développement embryonnaire et ont apporté les bases de l'embryologie moderne. C'est notamment Hans Driesch qui, en 1909, amène la théorie du développement à régulation à la suite de ses expériences sur les oursins de mer. Au début, il continue de croire que les cellules ont un destin fixe dans le rôle de la formation, mais qu'elles possèdent aussi cette capacité de régulation capable de contrer les possibles anormalités au moment du développement. Plus tard, par contre, il rejette toute idée de destin fixe et croit en la présence d'une capacité de totipotence chez toutes les cellules comprises dans le développement des embryons[2]. C'est ainsi que les deux idéologies embryologistes ont commencé à s'opposer: le développement à régulation surtout associé au clivage radial ou indéterminé et le développement en mosaïque surtout associé au clivage spiralé[3].
La première expérience menant au développement à régulation
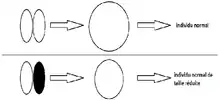
Travaillant à la station biologique de Naples, Hans Driesch décide de faire son expérience sur une espèce en abondance dans cette région, soit l'oursin de mer Echinus microtuberculatus. Contrairement à son prédécesseur Wilhelm Roux, un autre embryologiste qui travaillait sur des grenouilles, Hans possède une méthode efficace pour séparer les blastomères des œufs d'oursin. Elle consiste à agiter fortement l'eau dans laquelle se trouvent les œufs. La séparation étant possible, Hans Driesch s'intéresse alors à la première division cellulaire chez les œufs d'oursin de mer fertilisés. Agitant de nombreux œufs d'oursin de mer au premier stade de la division cellulaire, il sépare les blastomères en deux pour ensuite les placer dans des pétris différents. Comme la seule théorie de développement connue de l'époque était le développement mosaïque, soit la théorie selon laquelle les cellules des embryons ont un destin déterminé d'avance, Hans Driesch s'attendait à avoir une moitié d'oursin dans chaque pétri, car les deux cellules se seraient divisées, formant les parties respectives de l'oursin. Contrairement à ses attentes, il s'est retrouvé avec des blastomères qui s'étaient développés en embryons normalement formés sauf pour la taille de ceux-ci qui était diminuée de moitié par rapport à leur taille normale. Certains de ces embryons étaient rendus au stade de blastulation et au stade larvaire. C'est après avoir refait l'expérience à plusieurs reprises qu'Hans Driesch se distance des idées des autres embryologistes pour apporter sa théorie du développement à régulation[2] - [4].
Autres expérimentations scientifiques
- Hans Driesch n'a pas été le seul à faire des expériences sur le développement à régulation. En effet, l'expérience d'Hans Driesch a mené à de nombreuses autres expériences dont le but étaient de déterminer quelles espèces avaient un développement en mosaïque et celles ayant un développement à régulation. Ainsi, l'une de ces expériences a été celle de Barbara Conta Boyer en 1971 qui décide de travailler sur Chilidia groenlandica, un Acoele qui faisait partie de la classe des platyhelminthes à l'époque. Les Acoeles forment aujourd'hui partie d'une classe à part, mais cela n'affecte en rien la portée des travaux de Mme Boyer. Ayant un clivage spiralé, avec des duos de micromères plutôt que des quatuors comme le reste des plathelminthes, il s'agissait d'une espèce de prédilection pour vérifier le lien entre le clivage spiralé et le mosaïsme. Utilisant une méthode de délétion des cellules beaucoup plus précise que celle d'Hans Driesch, Barbara Conta Boyer procède aux délétions de cellules à différents moments de la division cellulaire à l'aide d'aiguilles de tungstène qui piquent à travers la capsule intacte des œufs. Ainsi ses expériences consistent en délétion d'une des deux cellules du premier stade de division cellulaire (1) comme l'a fait Hans Driesch sur ses embryons, d'une délétion des deux macromères du premier duo (1A et 1B) (2), du deuxième duo (2A et 2B)(3) et du troisième duo (3A et 3B)(4) et finalement la délétion de la moitié de l'embryon à un stade développé (5). Les cellules enlevées dans chaque expérience sont observables sur la figure à droite, et possèdent le même numéro que celui en gras. Ses expériences qui servaient à tester le développement en mosaïque changent rapidement de but après les premiers résultats et visent dorénavant à déterminer s'il y a des capacités régulatrices chez les embryons et si celles-ci se produisent seulement dans les premiers stages de la division cellulaire. Barbara se fonde donc sur 6 critères de différenciation pour déterminer si les embryons sont normaux et donc la présence de capacités régulatrices dans ses expériences. Les résultats obtenus dès le début viennent briser le concept selon lequel le clivage spiralé est symbole de mosaïsme. En effet, la première expérience montre qu'il y a une majorité d'individus normaux, résultat similaire à celui qu'a obtenu Hans Driesch avec ces oursins de mer. La deuxième expérience voit seulement 4 embryons éclore et aucun d'eux n’est normal. La troisième expérience et la quatrième expérience montrent chacun un embryon normal. La dernière expérience qui permettait de voir s'il y avait une capacité régulatrice des cellules dans les stages plus avancés de développement montre 26 individus normaux sur une possibilité de 33 sauf qu'ils avaient la moitié de la taille normale ce qui est dû au fait qu'ils ont été coupés en deux. Ainsi, La première et dernière expérience montrent qu'il y a des situations où la délétion cellulaire va être en mesure d'être régulée par le reste des cellules grâce à leur capacité totipotente et d'autres situations où la délétion cellulaire empêche le développement normal de l'individu comme dans les trois autres expériences qui ont obtenu des résultats négatifs lorsque les exceptions ne sont pas prises en compte. Cette expérience a donc prouvé que le développement à régulation est présent chez au moins une espèce à clivage spiralé, probablement parce qu'il s'agit d'une des espèces les plus primitives[3].
.png.webp) Figure montrant les différents stades de développement cellulaire lors de la division cellulaire et annotée conséquemment avec les délétions de l'expérience de Boyer en 1971.
Figure montrant les différents stades de développement cellulaire lors de la division cellulaire et annotée conséquemment avec les délétions de l'expérience de Boyer en 1971. - Quelques années plus tard, les chercheurs John B. Morrill, Claudia A. Blair et William J. Larsen du département de biologie de l'Université de Wesleyan se penchent sur les capacités régulatrices d'un gastropode, Lymnaea palustris lors de la délétion de cellules au stade 2,4,8 et 16 de la division cellulaire. Ils utilisent alors des aiguilles de verre ou l'agitation des œufs comme Hans Driesch pour faire les différentes délétions. Les résultats pour la délétion d'une cellule au premier stade de la division cellulaire sont encore une fois identiques aux résultats obtenus dans les expériences de développement à régulation précédente avec 39 des 444 œufs qui se sont développés normalement démontrant que Lymnaea palustris est un gastropode avec une capacité régulatrice, mais que celle-ci ne permet pas de sauver tous les individus. En effet, c'est moins de 9 % des œufs qui se sont développés normalement, un peu moins de 4 % avec la méthode de l'aiguille et un peu plus de 20 % avec la technique de l'agitation des œufs. Les résultats pour les autres délétions ont donné d'autres individus normaux pour le stade 4 de la division cellulaire. Il s'agit donc de la deuxième espèce avec clivage spiralé chez laquelle on a la preuve de capacité régulatrice symbole de développement à régulation[5].
Ces deux expériences montrent bien que le développement à régulation n'est pas encore compris complètement, et surtout qu'il semble être plus complexe que ce qu'Hans Driesch avait décrit.
Redéfinition du développement à régulation
La théorie du développement à régulation a été révisée à plusieurs reprises, notamment par Klaus Sander en 1971 afin d'y ajouter le fait que le concept de régulation devrait faire référence à la flexibilité de l'embryon, c'est-à-dire que lorsque celui-ci est soumis à des expérimentations, il peut également faire diverger le destin des cellules en éloignant de la normale ou en se rapprochant de la normale. C'est donc la capacité de régulation qui est importante, et non pas la direction dans laquelle la régulation se fait. Sander a également amené l'influence de l'interaction des cellules embryonnaires dans le processus de régulation, entraînant une nouvelles séries d'expériences. Les expériences ne portent donc plus sur la présence de la capacité régulatrice chez une espèce, mais plutôt sur les gènes et l'impact de leurs signaux dans les cellules embryonnaires pour permettre la régulation de l'espèce[1].
Le développement à régulation et la génétique
Avec la découverte des nombreuses interactions cellulaires, il est devenu impossible d'avoir un modèle embryonnaire de développement mosaïque ou de développement à régulation. Le développement à régulation continue à faire partie de la science qu'est l'évolution et le développement comme une trace du passé de l'embryologie, mais il est passé date et étudié sous une forme adaptée à la génétique moderne. En effet, on parle maintenant de patron de développement d'un individu. Bien sûr, il y a encore de la régulation, mais celle-ci est maintenant associée à la flexibilité des cellules dans le patron de développement. Ainsi certaines cellules possèdent des capacités régulatrices, mais ce ne sont jamais toutes les cellules. Les expériences visent maintenant à déterminer les gènes et les signaux impliqués dans cette régulation comme c'est le cas pour des protéines comme Hedgehog qui envoient des signaux aux cellules sur de longues distances ou bien des protéines qui envoient des signaux à faible portée visant à modifier la fonction de cellules voisines[1]. Le développement à régulation est donc une notion qui fait maintenant partie d'un ensemble de principes de développement découverts grâce à la génétique, qui aura tout de même eu un impact sur l'embryologie et fait avancer la science.
Voir aussi
Articles connexes
Lien externe
Notes et références
- Lawrence, Peter A., and Michael Levine. "Mosaic and regulative development: two faces of one coin." Current biology 16.7 (2006): R236-R239.
- Gilbert, Scott F., ed. A Conceptual History of Modern Embryology: Volume 7: A Conceptual History of Modern Embryology. Vol. 7. Springer Science & Business Media, 2013.
- Boyer, Barbara Conta. "Regulative development in a spiralian embryo as shown by cell deletion experiments on the acoel, Childia." Journal of Experimental Zoology 176.1 (1971): 97-105.
- (de) Hans Driesch, « Die isolirten Blastomeren des Echinidenkeimes », Archiv für Entwicklungsmechanik der Organismen, vol. 10, , p. 361-410 (ISSN 0365-6039 et 1432-041X, DOI 10.1007/BF02156642, lire en ligne, consulté le )
- Morrill, John B., Claudia A. Blair, and William J. Larsen. "Regulative development in the pulmonate gastropod, Lymnaea palustris, as determined by blastomere deletion experiments." Journal of Experimental Zoology 183.1 (1973): 47-55.